Мышиные модели метастазов рака груди - Mouse models of breast cancer metastasis
Мышиные модели с метастазами рака груди экспериментальные подходы, в которых мыши генетически модифицированный разработать опухоль молочной железы приводящие к отдаленным очаговым поражениям молочной железы эпителий сделано метастаз. Рак молочной железы у мышей может быть вызван генетическими мутации которые были идентифицированы при раке человека. Это означает, что модели могут быть созданы на основе молекулярных повреждений, соответствующих заболеванию человека.
Метастаз рака груди
Метастаз это процесс миграция из опухолевые клетки от первичного очага рака до удаленного места, где раковые клетки образуют вторичные опухоли. Метастатический рак молочной железы представляет собой наиболее разрушительный признак рака и считается заболеванием на поздней стадии.[1] Рак груди человека метастазирует в несколько отдаленных органов, таких как мозг, легкие, кости и печень.
Генетическое разнообразие между первичной и метастатической опухолью
Классическая теория, разработанная в начале 70-х годов, предполагала, что метастазирование происходит из-за генетически детерминированных субпопуляций первичных опухолей.[2] Генетический отклонение между метастатическими очагами имеет значение только для отдельных локус и внутри конкретных популяций клеток или только в одной клеточной популяции обнаруживаются различия, а некоторые локусы расходятся только в одной субпопуляции клеток. Это объясняет концепцию неоднородность опухоли и порядок генетических событий во время эволюция опухоли. Многие из генов, управляющих ростом на первичном участке, могут определять распространение и колонизация на эктопический сайт.[3][4][5] Рак груди считается генетически и клинически гетерогенным заболеванием, поскольку он отражает неоднородность нормальной ткани молочной железы в ее происхождении 17873350.[6] Должен произойти ряд дискретных генетических событий, чтобы позволить отдельным опухолевым клеткам, которые обладают способностью расти на эктопическом участке. Прогрессирование метастазов зависит от регулирования программ развития и событий окружающей среды.[7] Метастатический потенциал субпопуляций в клетках молочных желез мышей теперь рассматривается как относительно раннее событие, и распространение происходит одновременно с преинвазивными или микроинвазивными поражениями.[8][9] Генетические профили первичных и метастатических поражения в груди карциномы показать большую степень клональный уместность между поражениями.[10][11] Существуют различные закономерности распространенности генетических мутаций в геномах первичной опухоли молочной железы и ее метастазов.[12][13][14] Это также подтверждает генетическую гетерогенность между первичными новообразование больных раком груди и их метастазами.[15][16]
Гены, участвующие в органоспецифических метастазах
Рак молочной железы фенотипы периодически выражать гены в метастазах, которые необходимы при метастатическом процессе. Метастатическое разнообразие опосредуется активацией генов, которые действуют как сцепление с органо-специфическим ростом.[17] Рост поражений на эктопическом участке зависит от множества сложных взаимодействий между метастатическими клетками и хозяином. гомеостатический механизмы. Смертельный белок-белковые взаимодействия в месте метастазирования помогают выжить адаптированный клетки.[18]
Создание мышиных моделей рака груди
Целевое выражение онкогены в эпителиальных клетках молочной железы мышей - способ моделирования рака груди человека. Мутация или сверхэкспрессия онкогенов может контролироваться экспрессией в очень специфическом клеточном контексте, а не во всем организме. Другой способ моделирования рака груди человека - целевое ингибирование гена-супрессора опухоли.[19]
Мыши в генетических исследованиях
- В 1909 г. Кларенс К. Литтл выведен первый инбредный штамм, DBA (Разбавленная, коричневая не агути) мышь.
- В 1915 году Н. М. Холдейн идентифицировал первую связь у мышей между Альбинос разведение мышей и розовых глаз хромосома Семь.
- В 1921 г. C57BL стала одной из самых широко используемых мышей в генетике и первой штаммом, у которого был свой геном последовательный.
- В 1982 г. Пальмитер и Бринстер имплантировал чужой ген в оплодотворенная яйцеклетка, наконец генерируя первый трансгенный мыши генетически спроектированный для экспрессии доминирующих онкогенов.[20]
- В 1982 году стимуляция самовыражения от MMTV-LTR (вирус опухоли молочной железы мыши - длинный концевой повтор) было выполнено несколькими раундами беременность и кормление грудью оценить актуальность сотового протоонкоген, c-myc.[21]
Человек и мышь: геномное сравнение
Генетические исследования распространенных заболеваний человека имеют значительные ограничения с практической и этический причины.[22] Человек Сотовые линии может использоваться для моделирования болезни, но трудно изучить процессы на ткань уровне внутри органа или по всему телу. Мыши могут быть хорошим представителем болезней человека, потому что:[23]
- Есть близкое сходство физиология, развитие и клеточная биология между мышами и людьми.
- У людей и мышей около 30 000 генов, кодирующих белок. Количество мышиных генов без соответствующего человеческого гомолога составляет менее 1%.
- 90% геномов человека и мыши синтенический.
- 40% геномов человека и мыши могут быть выровнены по нуклеотид уровень.
- У мышей относительно короткие беременность периоды.
- Мышам требуется короткое время, чтобы достичь половой зрелости.
- У мышей большие размеры помета.
- Наличие сотен мутаций, затрагивающих практически все ткани и аспекты развития.
Мыши не могут быть идеальной моделью для рака груди. В основном это связано с отсутствием точности во многих моделях. Глядя на метастаз, трудно определить точное местоположение, а также его частоту. Другая проблема связана с подтипами эпителия и невозможностью целенаправленно воздействовать на них при нацеливании на мутацию. Примером этого может быть определение развития опухолей у мышей K14-Cre BRCA2. В стандартном случае удаление BRCA2 не приводило к опухолевому генезу, но если р53 был мутирован и инактивирован, то опухольгенез возник бы. Следовательно, окончательного ответа о происхождении опухоли нет из-за дополнительной мутации в p53.[24]
Клеточные линии метастатической карциномы молочной железы мыши
Различные клеточные линии карциномы молочной железы мыши, такие как 4T1[25] и TS / A, являются метастазами у сингенных иммунокомпетентных мышей и могут использоваться для идентификации генов и путей, участвующих в метастатическом процессе.[26]
Простые модели трансплантации опухолей
Трансплантация опухолевых клеток в иммунодефицитный мыши - это инструмент для изучения рака груди и его метастатических эффектов. Трансплантация происходит как аллотрансплантаты или же ксенографический трансплантаты.[27] Обычно клетки человека привит в ослабленном иммунитете мышиный получатель. Прививка клеток с помощью внутрипротоковой трансплантации,[28] инъекциями очищенных жировых отложений молочной железы[29][30] или путем трансплантации в хвостовую вену.[31][32][33] Различные органы могут быть засеяны клетками рака груди в зависимости от пути введения.[34]
- Сердечная инъекция: кость
- Инъекция в хвостовую вену: легкое
- Инъекция в селезенку: печень
- Инъекция сонной артерии: мозг
Модели трансплантата опухолевой ткани
Использовали мышей с специфическим иммунодефицитом, мышей NOD / SCID (диабет без ожирения / тяжелый условный иммунодефицит). Эти мутации позволяют интегрировать новую ткань ксенотрансплантата. Мыши сначала должны гуманизировать жировые подушечки молочной железы путем инъекции иммортализованных телеморазой человеческих стромальных фибробластов молочной железы (фибробласты RMF / EG) в жировые подушечки молочных желез. Без этой инъекции эпителиальные клетки молочной железы человека, привитые к подушечке, не могут колонизировать и расти. Затем необходимо облучить фибробласт RMF / EG, чтобы обеспечить экспрессию ключевых белков и факторов роста. После 4 недель развития недавно привитые эпителиальные клетки молочной железы человека увеличивались в жировой подушке.[35]
Генно-инженерные мыши для изучения метастазов
Генно-инженерные мыши созданы для моделирования человека. фенотипы и патологии. Мутантные мыши могут включать: трансгены используя разные способы доставки:
- Использование бактериальной системы, индуцируемой тетрациклином, позволяющей включать и выключать (Система Tet-On / Tet-Off)[36]
- Целевые мутации стук в гене и последовательность выбивания используя Cre-Lox рекомбинация система[37]
- Представление о ретро вирусный мутации[38]
- Введение химически индуцированных мутаций
Трансгенные мышиные модели рака груди
Мыши, переживающие процесс трансгенеза, известны как трансгенные мыши. Базовый трансген имеет промоутер область, кодирующая последовательность белка, Интрон и стоп-кодон. Вирус опухоли молочной железы мышей (MMTV) - это ретро-вирус, который, как известно, вызывает опухоли груди после активации.[39] MMTV - это наследственный соматический мутаген, диапазон мишеней которого ограничен. Он содержит регуляторную последовательность ДНК, называемую длинным концевым повтором (LTR), которая способствует транскрипции, индуцируемой стероидными гормонами.[40][41] Опухоль, индуцированная вирусом опухоли молочной железы мыши, также может быть осуществлена путем интеграции вирусного генома. Известно, что сайты интеграции являются критическими генами для клеточной регуляции.[42]Сывороточный кислый белок (WAP),[43] - еще один распространенный промотор, используемый для создания моделей рака молочной железы у мышей. Список других промоторов, специфичных для молочных желез, и моделей мышей см.[44]
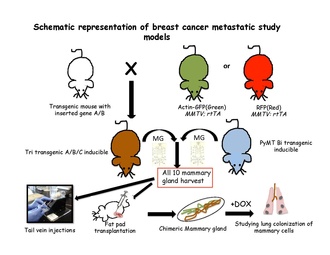
ММТВ-ПыМТ
MMTV-PyMT - это модель метастазирования рака груди, в которой MMTV-LTR используется для управления экспрессией специфических для молочной железы полиомавирус средний Т-антиген, что приводит к быстрому развитию высокометастатических опухолей.[45] MMTV-PyMT - наиболее часто используемая модель для изучения прогрессирования и метастазирования опухоли молочной железы. Затем мышей MMTV-PyMT скрещивают с другими генетически модифицированными мышами для создания различных типов моделей рака молочной железы, включая:
- Сигнализация PI3K / Akt при метастазировании может быть продемонстрировано в MMTV-PyMT; Akt1 - / - мыши.[46]
- Chemoattractive паракринная петля из колониестимулирующий фактор-1 (CSF-1) и EGF лиганды между опухоль-ассоциированными макрофагами (ТАМ) и опухолевыми клетками, а также метастазы в легкие могут быть изучены путем скрещивания мышей MMTV-PyMT с мышами Csf-1 - / -.[47]
- Роль врожденный и адаптивный иммунитет ответ на помощь при метастазировании можно изучить в MMTV-PyMT; Rag1 - / - мыши, у которых CD4 + Т-клетки выборочно теряются. Интерлейкин-4 (IL4) отсутствие модели ММТВ-ПыМТ; IL4 - / - мыши.[48]
- Роль молекулы адгезии CD44 при метастазах в легкие.[49]
- Условная абляция в клетках рака молочной железы MMTV-PyMT было сделано для выявления прометастатического действия ангиогенный факторы, Фактор роста эндотелия сосудов А (VEGF-A).[50]
- Роль автокринный трансформирующий фактор роста передача сигналов бета 1 (TGF-β1) на подвижность и выживаемость в клетках PymT, полученных из рака молочной железы мыши MMTV-PymT.[51]
- Остальные - MMTV-PyMT; uPA - / -[52] и ММТВ-ПыМТ; МЕКК1 - / -.[53]
ММТВ-HER2 / neu
MMTV-LTR может также использоваться для стимуляции рецепторной тирозин-протеинкиназы ErbB2 для трансформации эпителия молочной железы мыши. ErbB2 - это онкоген, усиленный и сверхэкспрессируемый примерно в 20% случаев рака груди человека. У мышей, несущих этот онкоген, примерно через 15 недель после беременности развиваются мультифокальные аденокарциномы с метастазами в легких.[54][55]Чтобы создать более точное представление о мутациях гена HER2, исследователи объединили ген мыши, содержащий neu, и ген крысы, содержащий neu. Это решает проблему с точки зрения моделирования амплификации HER2 в развитии мышей. У неслитых мышей молочная железа вернется к почти девственной форме, но с этим добавлением молочная железа сохранит развитую функцию.[56]
Би-трансгенные модели
Мышиные модели, имеющие два трансгена, называются би-трансгенными. Чтобы проверить взаимодействие двух онкогенов, Тим Стеверт и его группа в 1987 году создали первые би-трансгенные мышиные модели MMTV-Мой с и ММТВ- Рас скрещивание мышей привело к ускорению онкогенеза.[57] Выражение TGFβ в клетках рака груди MMTV-ErbB2; Двойные трансгенные мыши MMTV-TGFβ могут индуцировать более высокие уровни циркулирующих опухолевых клеток и метастазирование в легкие.[58] Ген Ras можно комбинировать с rtTA (трансактиватор обратного тетрациклина) для создания би-трансгенной индуцибельной модели мыши посредством контролируемой тетрациклином активации транскрипции, например мыши, несущие TetO-KrasG12D (TOR) и MMTV-rtTA (MTB), имеют трансген, экспрессирующий обратный трансактиватор тетрациклина (rtTA) в эпителиальных клетках молочных желез.[59]
Три-трансгенные модели
Три-трансгенные мышиные модели состоят из более чем двух генов. Множественные комбинации и генетические модификации выполняются таким образом, что один или все гены переводятся в состояние непрерывной экспрессии или контролируемым образом для их активации в разные моменты времени. Например, ТОМ (TetO-myc); ТЗ; Мыши MTB, у которых гены myc (M) и ras (R) находятся под контролем операторов тетрациклина. Их также можно как активировать, так и деактивировать, добавляя доксициклин. Другие комбинации в этом отношении: TOM; Крас; MTB, где myc может индуцироваться и не индуцироваться в различные моменты времени, пока Kras находится в непрерывно экспрессируемом состоянии, и myc; ТЗ; Модель МТБ наоборот.[60]
Применение генетически модифицированных мышей для изучения метастазов
Метастатический каскад можно изучить, удерживая активацию гена под контролем или добавляя репортерный ген, например. Бета-актин GFP (зеленый флуоресцентный белок) или RFP (красный флуоресцентный белок).
Идентификация генов, регулирующих метастазирование
Путем включения / выключения определенных генов путем гомологичной рекомбинации можно измерить степень метастазирования и, например, достичь идентификации новых генов-мишеней. геном, который последовательно регулирует метастатическое поведение раковых клеток, является TGF-β1. Резкое прекращение передачи сигналов TGF-β в опухолевых клетках молочной железы MMTV-PyMT приводит к пятикратному увеличению метастазов в легких.[61]Некоторые энхансерные области также могут быть проанализированы и могут быть определены как критически важные части пролиферации клеток, например усиливающая область, связанная с критическим для рака геном p53, который был определен с помощью CRISPR-Cas9.[62]
Отслеживание происхождения в моделях метастазов
Стратегии количественного отслеживания клонов доказали свою эффективность в разрешении судьбы клеток в нормальных эпителиальных тканях либо с использованием тканеспецифических, либо стволовая клетка -специфические трансгены. Для проведения эксперимента по индуцибельному отслеживанию клонов в геном мыши должны быть встроены два компонента: переключатель и репортер. Переключатель обычно представляет собой регулируемую лекарственными средствами форму бактериального фермента Cre-рекомбиназы. Этот фермент распознает определенные последовательности, называемые сайтами LoxP.[63] Белки, способные усилить идентификацию меченых клеток или конкретной популяции немеченых клеток, кодируются репортерными трансгенами. После взятия всех десяти молочных желез мышей у трансгенных мышей обычно готовят суспензию единичных клеток и трансплантируют либо в хвостовую вену нетрансгенных мышей-реципиентов.[31] или в очищенной жировой подушке нетрансгенных мышей, повторно заселяющих жировую подушку молочной железы.[64] Затем эти клетки отслеживаются в кровотоке, легких, костном мозге и печени, чтобы найти благоприятное место для метастазирования. Эти трансгенные клетки можно проследить в соответствии с их особенностями флуоресценции или индуцирования путем помещения реципиентов на доксициклин.
Циркулирующие опухолевые клетки
Еще один инструмент для изучения метастазов рака молочной железы - это поиск циркулирующих опухолевых клеток у трансгенных мышей, например. Мыши MMTV-PyMT могут реагировать на различные методы лечения, выделяя опухолевые клетки в кровь, что приводит к метастазам в легких.[65] Не только в крови, но и в костном мозге можно обнаружить клетки, например цитокератин -положительные клетки в костном мозге трансгенных мышей MMTV-pyMT и MMTV-Neu были идентифицированы, но не в контроле дикого типа.[66]
Ограничения
В отсутствие специфических маркеров для клеток молочной железы модели с генетической маркировкой опухолевых клеток дают лучшее экспериментальное преимущество, однако небольшой объем периферической крови, который можно получить от живых животных, ограничивает применение этого метода.
В естественных условиях визуализация метастатических моделей мышей
Модели трансгенных мышей можно визуализировать с помощью различных неинвазивных методов.
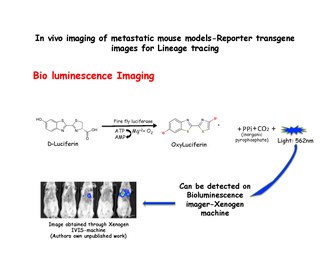
Биолюминесцентная визуализация
Биолюминесцентная визуализация основан на обнаружении света, производимого ферментативным окислением экзогенного субстрата. Субстрат люциферин окисляется до оксилюциферина в присутствии люцифераза и излучает свет, который можно обнаружить с помощью системы IVIS, такой как аппарат Xenogen. Диссоциированные клетки молочной железы из MMTV-PyMT: IRES: Luc; MTB (Внутренний сайт входа рибосомы: Люциферин ) животным (которые не подвергались воздействию доксициклина) можно вводить инъекции в боковые хвостовые вены мышей с иммунодефицитом на диете без доксициклина. Сигнал биолюминесценции не будет наблюдаться в легких мышей-реципиентов до тех пор, пока им не будет дан корм с доксициклином. Затем биолюминесценция может быть обнаружена в груди в течение 2 недель после начала воздействия доксициклина.[31] Люцифераза вводится непосредственно перед съемкой изображений.
Флуоресцентная визуализация
Прижизненная микроскопия с многофотонным возбуждением - это метод визуализации генно-инженерных клеток непосредственно in vivo. Многоступенчатые метастатические каскады могут быть визуализированы путем маркировки уникальным флуоресцентным цветом под флуоресцентный микроскоп.[67][68]
Радиоизотопные изображения
Позитронно-эмиссионная томография (ДОМАШНИЙ ПИТОМЕЦ), однофотонная эмиссионная компьютерная томография (ОФЭКТ) и компьютерная томография (КТ) использовались для сравнения эффективности этих изображений in vivo для обнаружения поражений на ранней стадии и для оценки ответа на химиотерапию.[69]
МРТ
Магнитно-резонансная томография требует использования наночастиц (липосом) и контрастного вещества для МРТ, называемого гадолинием. Затем частицы помещали в везикулы через фильтр с поликарбонатной мембраной. Наночастицы вводят в образующихся метастазов мышей и оставляют там на двадцать четыре часа. Затем этих мышей сканируют, и в программном обеспечении для визуализации обнаруживаются скопления этих частиц в определенных областях, где клетки метастазировали.[22]
Смотрите также
- Ансамбль база данных генома модельных организмов
- Карта судьбы
- Люциферин светлячка
- Нацеливание на гены
- Захват генов
- Генетическая рекомбинация
- История модельных организмов
- Гомологичная рекомбинация
- Обмен кассет, опосредованный рекомбиназой
- Технология сайт-специфической рекомбиназы
Рекомендации
- ^ Hanahan, D .; Вайнберг, Р. (2000). «Признаки рака». Клетка. 100 (1): 57–70. Дои:10.1016 / S0092-8674 (00) 81683-9. PMID 10647931.
- ^ Фидлер, И. Дж. (1973). «Выбор последовательных линий опухоли для метастазирования». Природа Новая Биология. 242 (118): 148–9. Дои:10.1038 / newbio242148a0. PMID 4512654.
- ^ Martins, F.C .; Де, S; Альмендро, V; Gönen, M; Park, S. Y .; Blum, J. L .; Херлихи, W; Ethington, G; Schnitt, S.J .; Тунг, Н; Гарбер, Дж. Э .; Феттен, К; Michor, F; Поляк, К (2012). «Эволюционные пути в опухолях молочной железы, связанных с BRCA1». Открытие рака. 2 (6): 503–11. Дои:10.1158 / 2159-8290.CD-11-0325. ЧВК 3738298. PMID 22628410.
- ^ Shah, S.P .; Morin, R.D .; Khattra, J; Прентис, L; Пью, Т; Берли, А; Делани, А; Гельмон, К; Гулианы, Р; Senz, J; Steidl, C; Holt, R.A .; Джонс, S; Вс, М; Leung, G; Мур, Р.; Северсон, Т; Тейлор, Г. А .; Teschendorff, A.E .; Це, К; Турашвили, Г; Varhol, R; Warren, R.L .; Watson, P; Чжао, Y; Caldas, C; Хантсман, Д; Херст, М; Marra, M. A .; Aparicio, S (2009). «Мутационная эволюция в дольчатой опухоли молочной железы, профилированная при разрешении одного нуклеотида». Природа. 461 (7265): 809–13. Дои:10.1038 / природа08489. PMID 19812674.
- ^ Geyer, F.C .; Вайгельт, В; Натраджан, Р. Lambros, M. B .; De Biase, D; Вачева, Р; Сэвидж, К; Маккей, А; Ашворт, А; Рейс-Филхо, Дж. С. (2010). «Молекулярный анализ показывает генетическую основу фенотипического разнообразия метапластических карцином груди». Журнал патологии. 220 (5): 562–73. Дои:10.1002 / путь.2675. PMID 20099298.
- ^ Ашкенази, Р. Jackson, T. L .; Dontu, G; Вича, М. С. (2007). «Возможности исследования стволовых клеток рака груди с использованием математического моделирования». Стволовые клетки. 3 (2): 176–82. Дои:10.1007 / s12015-007-0026-2. PMID 17873350.
- ^ Мюллер, А; Хоми, B; Сото, Н; Ge, N; Катрон, Д; Бьюкенен, М. Э .; Маккланахан, Т; Мерфи, Э; Юань, Вт; Вагнер, С. Н .; Barrera, J. L .; Мохар, А; Verástegui, E; Злотник, А (2001). «Участие хемокиновых рецепторов в метастазировании рака груди». Природа. 410 (6824): 50–6. Дои:10.1038/35065016. PMID 11242036.
- ^ Кляйн, К. А. (2009). «Параллельное развитие первичных опухолей и метастазов». Обзоры природы Рак. 9 (4): 302–12. Дои:10.1038 / nrc2627. PMID 19308069.
- ^ Weng, D; Penzner, J. H .; Песня, B; Koido, S; Calderwood, S.K .; Гонг, Дж (2012). «Метастазирование является ранним событием при карциноме молочной железы у мышей и связано с клетками, несущими маркеры стволовых клеток». Исследование рака груди. 14 (1): R18. Дои:10.1186 / bcr3102. ЧВК 3496135. PMID 22277639.
- ^ Лю, Вт; Laitinen, S; Хан, S; Вихинен, М; Ковальски, Дж; Ю, Г; Чен, L; Ewing, C.M .; Eisenberger, M. A .; Кардуччи, М. А .; Nelson, W.G .; Егнасубраманян, S; Луо, Дж; Ван, Y; Сюй, Дж; Isaacs, W. B .; Visakorpi, T; Бова, Г. С. (2009). «Анализ числа копий указывает на моноклональное происхождение летального метастатического рака простаты». Природа Медицина. 15 (5): 559–65. Дои:10,1038 / нм.1944. ЧВК 2839160. PMID 19363497.
- ^ Торрес, L; Ribeiro, F. R .; Пандис, Н. Андерсен, Дж. А .; Heim, S; Тейшейра, М. Р. (2007). «Внутриопухолевая геномная гетерогенность рака молочной железы с клональной дивергенцией между первичными карциномами и метастазами в лимфатические узлы». Исследования и лечение рака груди. 102 (2): 143–55. Дои:10.1007 / s10549-006-9317-6. PMID 16906480.
- ^ Пандис, Н. Teixeira, M. R .; Адейинка, А; Ризу, H; Барди, G; Mertens, F; Андерсен, Дж. А .; Бондесон, L; Сфикас, К; Qvist, H; Апостоликас, N; Мительман, Ф; Хайм, S (1998). «Цитогенетическое сравнение первичных опухолей и метастазов в лимфатические узлы у больных раком груди». Гены, хромосомы и рак. 22 (2): 122–9. Дои:10.1002 / (SICI) 1098-2264 (199806) 22: 2 <122 :: AID-GCC6> 3.0.CO; 2-Z. PMID 9598799.
- ^ Куукасярви, Т; Karhu, R; Таннер, М; Кяхконен, М; Schäffer, A; Nupponen, N; Pennanen, S; Каллиониеми, А; Kallioniemi, O.P .; Изола, Дж. (1997). «Генетическая гетерогенность и клональная эволюция, лежащие в основе развития асинхронных метастазов при раке груди человека». Исследования рака. 57 (8): 1597–604. PMID 9108466.
- ^ Bonsing, B.A .; Corver, W. E .; Fleuren, G.J .; Cleton-Jansen, A.M .; Девили, П; Корнелисс, К. Дж. (2000). «Аллелотипический анализ распределенных по потоку клеток рака молочной железы демонстрирует генетически связанные диплоидные и анеуплоидные субпопуляции в первичных опухолях и метастазах в лимфатические узлы». Гены, хромосомы и рак. 28 (2): 173–83. Дои:10.1002 / (SICI) 1098-2264 (200006) 28: 2 <173 :: AID-GCC6> 3.0.CO; 2-1. PMID 10825002.
- ^ Wu, J. M .; Fackler, M. J .; Галушка, М.К .; Molavi, D.W .; Тейлор, М. Э .; Teo, W. W .; Гриффин, К; Феттинг, Дж; Дэвидсон, Н.Е .; Де Марсо, А. М .; Hicks, J. L .; Chitale, D; Ladanyi, M; Сукумар, S; Аргани, П. (2008). «Гетерогенность метастазов рака молочной железы: сравнение терапевтической целевой экспрессии и метилирования промотора между первичными опухолями и их мультифокальными метастазами». Клинические исследования рака. 14 (7): 1938–46. Дои:10.1158 / 1078-0432.CCR-07-4082. ЧВК 2965068. PMID 18381931.
- ^ Шмидт-Киттлер, О; Рэгг, Т; Даскалакис, А; Granzow, M; Ahr, A; Бланкенштейн, Т. Дж .; Кауфманн, М; Диболд, Дж; Арнхольдт, Н; Muller, P; Бишофф, Дж; Харич, Д; Шлимок, Г; Ритмюллер, G; Эйлс, Р. Кляйн, К. А. (2003). «От скрытых диссеминированных клеток к явным метастазам: генетический анализ прогрессирования системного рака молочной железы». Труды Национальной академии наук. 100 (13): 7737–42. Дои:10.1073 / pnas.1331931100. ЧВК 164657. PMID 12808139.
- ^ Мартин, B; Sanz, R; Aragüés, R; Олива, Б; Сьерра, А (2008). «Функциональная кластеризация белков метастазов описывает ресурсы пластической адаптации клеток рака груди к новым микросредам». Журнал протеомных исследований. 7 (8): 3242–53. Дои:10.1021 / pr800137w. PMID 18582095.
- ^ Langley, R. R .; Фидлер, И. Дж. (2007). «Взаимодействие опухолевой клетки и микроокружения в патогенезе метастазирования рака». Эндокринные обзоры. 28 (3): 297–321. Дои:10.1210 / er.2006-0027. PMID 17409287.
- ^ Гупта, ПБ; Купервассер, К. (2004). Модели заболеваний рака груди. Открытие лекарств сегодня: модели болезней 1 (1), 9-16. DOI: 10.1016 / j.ddmod.2004.05.001
- ^ Palmiter, R.D .; Brinster, R.L .; Hammer, R.E .; Trumbauer, M.E .; Розенфельд, М. Г .; Birnberg, N.C .; Эванс, Р. М. (1992). «Резкий рост мышей, которые развиваются из яиц, в которые вводят слитые гены металлотионеина и гормона роста. 1982». Биотехнология (Ридинг, Массачусетс). 24: 429–33. PMID 1422050.
- ^ Nusse, R; Вармус, Х. Э. (1982). «Многие опухоли, вызванные вирусом опухоли молочной железы мыши, содержат провирус, интегрированный в одну и ту же область генома хозяина». Клетка. 31 (1): 99–109. Дои:10.1016/0092-8674(82)90409-3. PMID 6297757.
- ^ а б Goldman, E; Зингер, А; Сильва, ДД; Yaari, Z; Варди-Окнин, Д; Гольдфедер, М; Schroeder, JE; Шаинский-Ройтман, Дж; Гершковиц, Д; Шредер, А; (2017). Наночастицы нацелены на метастазы рака груди на ранней стадии in vivo. Нанотехнологии 28 (43), 1-13. DOI: 10.1086 / 13616528 / aa8a3d
- ^ Chinwalla, A. T .; Waterston, L.L .; Lindblad-Toh, K.D .; Birney, G.A .; Роджерс, Л. А .; Abril, R. S .; Agarwal, T. A .; Agarwala, L.W .; Ainscough, E. R .; Alexandersson, J. D .; An, T. L .; Antonarakis, W. E .; Attwood, J. O .; Baertsch, M.N .; Bailey, K. H .; Barlow, C. S .; Beck, T. C .; Берри, В .; Birren, J .; Bloom, E .; Bork, R.H .; Botcherby, M. C .; Bray, R.K .; Brent, S.P .; Brown, P .; Brown, E .; Bult, B .; Burton, T .; Батлер, Д.Г .; и другие. (2002). «Первоначальное секвенирование и сравнительный анализ генома мыши». Природа. 420 (6915): 520–562. Дои:10.1038 / природа01262. PMID 12466850.
- ^ Вагнер, К.В. (2003). Модели рака груди: quo vadis, моделирование животных? Исследование рака груди 6 (31), 31-38.doi: 10.1186 / bcr723
- ^ Пуласки Б.А., С. Остранд-Розенберг. 2001. "Модель опухоли молочной железы мыши 4T1". Curr Protoc Immunol. Глава 20: Раздел 20.2. DOI: 10.1002 / 0471142735.im2002s39
- ^ Knott SRV, E Wagenblast, S Khan, SY Kim, M Soto, M Wagner, MO Turgeon, L Fish, N Erard, AL Gable, AR Maceli, S Dickopf, EK Papachristou, CS D'Santos, LA Carey, JE Wilkinson, JC Harrell, CM Perou, H Goodarzi, G Poulogiannis и GJ Hannon. 2018. «Биодоступность аспарагина определяет метастазирование в модели рака груди». Природа. DOI: 10.1038 / природа25465
- ^ Ханна, К; Хантер, К. (2005). «Моделирование метастазов in vivo». Канцерогенез. 26 (3): 513–23. Дои:10.1093 / carcin / bgh261. PMID 15358632.
- ^ Бехбод, Ф; Kittrell, F. S .; Ламарка, Н; Эдвардс, D; Кербави, S; Heestand, J.C .; Янг, E; Mukhopadhyay, P; Yeh, H.W .; Allred, D.C .; Ху, М; Поляк, К; Rosen, J.M .; Медина, Д. (2009). «Модель внутрипротоковой трансплантации человек-мышь имитирует подтипы протоковой карциномы in situ». Исследование рака груди. 11 (5): R66. Дои:10.1186 / bcr2358. ЧВК 2790841. PMID 19735549.
- ^ Aslakson, C.J .; Миллер, Ф. Р. (1992). «Избирательные события в метастатическом процессе, определенные анализом последовательного распространения субпопуляций опухоли молочной железы мыши». Исследования рака. 52 (6): 1399–405. PMID 1540948.
- ^ Ян, Цзин; Мани, Сендурай А; Донахер, Джоана Лю; Рамасвами, Шридхар; Ициксон, Рафаэль А; Пойдем, Кристоф; Савагнер, Пьер; Гительман, Инна; Ричардсон, Андреа; Вайнберг, Роберт А. (2004). «Твист, главный регулятор морфогенеза, играет важную роль в метастазировании опухолей». Клетка. 117 (7): 927–39. Дои:10.1016 / j.cell.2004.06.006. PMID 15210113.
- ^ а б c Подсыпанина, К; Du, Y. C .; Jechlinger, M; Беверли, Л. Дж .; Амбарцумян, Д; Вармус, Х (2008). «Посев и распространение нетрансформированных клеток молочной железы мыши в легких». Наука. 321 (5897): 1841–4. Дои:10.1126 / science.1161621. ЧВК 2694414. PMID 18755941.
- ^ Талмадж, Дж. Э .; Сингх, Р. К .; Fidler, I.J .; Раз, А (2007). «Мышиные модели для оценки новых и традиционных терапевтических стратегий лечения рака». Американский журнал патологии. 170 (3): 793–804. Дои:10.2353 / ajpath.2007.060929. ЧВК 1864878. PMID 17322365.
- ^ Kim, M. Y .; Оскарссон, Т; Ачарья, S; Nguyen, D. X .; Zhang, X. H .; Нортон, L; Massagué, J (2009). «Самосев опухоли циркулирующими раковыми клетками». Клетка. 139 (7): 1315–26. Дои:10.1016 / j.cell.2009.11.025. ЧВК 2810531. PMID 20064377.
- ^ Варго-Гогола, Т; Розен, Дж. М. (2007). «Моделирование рака груди: один размер не подходит всем». Обзоры природы Рак. 7 (9): 659–72. Дои:10.1038 / nrc2193. PMID 17721431.
- ^ Купервассер, К; Чаваррия, Т; Ву, М; Magrane, G; Грей, JW; Кэри, L; Ричардсон, А; Вайнберг, РА. (2004). Реконструкция функционально нормальной и злокачественной ткани груди человека у мышей. Пнас 101 (14), 4966-4971. DOI: 10.1073 / pnas.0401064101
- ^ Госсен, М; Bujard, H (1992). «Жесткий контроль экспрессии генов в клетках млекопитающих с помощью промоторов, чувствительных к тетрациклину». Труды Национальной академии наук Соединенных Штатов Америки. 89 (12): 5547–51. Дои:10.1073 / пнас.89.12.5547. ЧВК 49329. PMID 1319065.
- ^ Зауэр, В; Хендерсон, Н. (1989). «Cre-стимулированная рекомбинация на loxP-содержащих последовательностях ДНК, помещенных в геном млекопитающих». Исследования нуклеиновых кислот. 17 (1): 147–61. Дои:10.1093 / nar / 17.1.147. ЧВК 331541. PMID 2783482.
- ^ Ду, З; Подсыпанина, К; Хуанг, S; МакГрат, А; Toneff, M. J .; Богословская, Е; Чжан, X; Moraes, R.C .; Флак, М; Allred, D.C .; Льюис, М. Т .; Varmus, H.E .; Ли, Y (2006). «Введение онкогенов в молочные железы in vivo с помощью птичьего ретровирусного вектора инициирует и способствует канцерогенезу на моделях мышей». Труды Национальной академии наук. 103 (46): 17396–401. Дои:10.1073 / pnas.0608607103. ЧВК 1635021. PMID 17090666.
- ^ Каллахан, Р. Смит, Г. Х. (2000). «MMTV-индуцированный туморогенез молочной железы: открытие генов, развитие злокачественных новообразований и клеточные пути». Онкоген. 19 (8): 992–1001. Дои:10.1038 / sj.onc.1203276. PMID 10713682.
- ^ Ringold, G.M .; Ямамото, К. Р .; Tomkins, G.M .; Бишоп, М. Вармус, Х. Э. (1975). «Дексаметазон-опосредованная индукция РНК вируса опухоли молочной железы мышей: система для изучения действия глюкокортикоидов». Клетка. 6 (3): 299–305. Дои:10.1016/0092-8674(75)90181-6. PMID 212202.
- ^ Ямамото, К. Р .; Пайвар, Ф; Firestone, G.L .; Maler, B.A .; Рэндж, О; Карлштедт-Дюк, Дж; Gustafsson, J. A .; Чендлер, В. Л. (1983). «Биологическая активность клонированных фрагментов ДНК вируса опухоли молочной железы, которые связывают очищенный белок глюкокортикоидного рецептора in vitro». Симпозиумы Колд-Спринг-Харбор по количественной биологии. 47 (2): 977–84. Дои:10.1101 / sqb.1983.047.01.111. PMID 6305596.
- ^ Росс, RS. (2010). Молекулярная биология и онкогенез вируса опухоли молочной железы мышей. Вирусы 2 (9), 2000-2012 гг. DOI: 10.3390 / v2092000
- ^ Кэмпбелл, С. М .; Rosen, J.M .; Hennighausen, L.G .; Strech-Jurk, U; Сиппель, А. Э. (1984). «Сравнение генов кислого белка сыворотки крысы и мыши». Исследования нуклеиновых кислот. 12 (22): 8685–97. Дои:10.1093 / nar / 12.22.8685. ЧВК 320407. PMID 6095207.
- ^ Фантоцци, А; Кристофори, Г. (2006). «Мышиные модели метастазов рака груди». Исследование рака груди. 8 (4): 212. Дои:10.1186 / bcr1530. ЧВК 1779475. PMID 16887003.
- ^ Guy, C.T .; Кардифф, Р. Д .; Мюллер, В. Дж. (1992). «Индукция опухолей молочной железы путем экспрессии онкогена среднего Т полиомавируса: модель трансгенных мышей для метастатического заболевания». Молекулярная и клеточная биология. 12 (3): 954–61. Дои:10.1128 / mcb.12.3.954. ЧВК 369527. PMID 1312220.
- ^ Кларенбек, S; Ван Мильтенбург, М. Х .; Йонкерс, Дж (2013). «Генетически сконструированные мышиные модели передачи сигналов PI3K при раке груди». Молекулярная онкология. 7 (2): 146–64. Дои:10.1016 / j.molonc.2013.02.003. ЧВК 5528412. PMID 23478237.
- ^ Lin, E. Y .; Nguyen, A. V .; Russell, R.G .; Поллард, Дж. У. (2001). «Колониестимулирующий фактор 1 способствует прогрессированию опухолей молочной железы до злокачественных новообразований». Журнал экспериментальной медицины. 193 (6): 727–40. Дои:10.1084 / jem.193.6.727. ЧВК 2193412. PMID 11257139.
- ^ Denardo, D.G .; Barreto, J. B .; Андреу, П; Васкес, L; Tawfik, D; Колхаткар, Н; Куссенс, Л. М. (2009). «CD4 (+) Т-клетки регулируют легочные метастазы карциномы молочной железы, усиливая протопухолевые свойства макрофагов». Раковая клетка. 16 (2): 91–102. Дои:10.1016 / j.ccr.2009.06.018. ЧВК 2778576. PMID 19647220.
- ^ Lopez, J. I .; Camenisch, T. D .; Стивенс, М. В .; Sands, B.J .; Макдональд, Дж; Шредер, Дж. А. (2005). «CD44 ослабляет метастатическую инвазию во время прогрессирования рака груди». Исследования рака. 65 (15): 6755–63. Дои:10.1158 / 0008-5472.CAN-05-0863. PMID 16061657.
- ^ Schoeffner, D. J .; Matheny, S. L .; Акахане, Т; Фактор, В; Берри, А; Мерлино, G; Торгейрссон, У. П. (2005). «VEGF способствует росту опухоли молочной железы у трансгенных мышей через паракринные и аутокринные механизмы». Лабораторные исследования. 85 (5): 608–23. Дои:10.1038 / labinvest.3700258. PMID 15765121.
- ^ Muraoka-Cook, R.S .; Курокава, H; Koh, Y; Forbes, J. T .; Roebuck, L.R .; Barcellos-Hoff, M.H .; Moody, S.E .; Chodosh, L.A .; Артеага, К. Л. (2004). «Условная сверхэкспрессия активного трансформирующего фактора роста бета1 in vivo ускоряет метастазирование трансгенных опухолей молочной железы». Исследования рака. 64 (24): 9002–11. Дои:10.1158 / 0008-5472.CAN-04-2111. PMID 15604265.
- ^ Альмхольт, К; Lund, L.R .; Rygaard, J; Nielsen, B.S .; Данё, К; Рёмер, Дж; Йонсен, М. (2005). «Уменьшение метастазирования трансгенного рака молочной железы у мышей с дефицитом урокиназы». Международный журнал рака. 113 (4): 525–32. Дои:10.1002 / ijc.20631. PMID 15472905.
- ^ Cuevas, B.D .; Винтер-Ванн, А. М .; Johnson, N.L .; Джонсон, Г. Л. (2006). «MEKK1 контролирует деградацию матрикса и диссеминацию опухолевых клеток во время метастазирования полиомы среднего Т-управляемого рака молочной железы». Онкоген. 25 (36): 4998–5010. Дои:10.1038 / sj.onc.1209507. PMID 16568086.
- ^ Slamon, D. J .; Clark, G.M .; Wong, S.G .; Levin, W. J .; Ульрих, А; Макгуайр, В. Л. (1987). «Рак груди человека: корреляция рецидива и выживаемости с амплификацией онкогена HER-2 / neu». Наука. 235 (4785): 177–82. Дои:10.1126 / science.3798106. PMID 3798106.
- ^ Muller, W. J .; Sinn, E; Pattengale, P.K .; Уоллес, Р. Ледер, П. (1988). «Одностадийная индукция аденокарциномы молочной железы у трансгенных мышей, несущих активированный онкоген c-neu». Клетка. 54 (1): 105–15. Дои:10.1016/0092-8674(88)90184-5. PMID 2898299.
- ^ Фрай, EA; Taneka, P; Иноуэ, К. (2016). Онкогенные и опухолевые мышиные модели рака груди, использующие HER2 / neu. Международный журнал рака 140 (3), 495-503. DOI: 10.1002 / ijc.30399
- ^ Sinn, E; Мюллер, Вт; Pattengale, P; Теплер, I; Уоллес, Р. Ледер, П. (1987). "Коэкспрессия генов MMTV / v-Ha-ras и MMTV / c-myc у трансгенных мышей: синергетическое действие онкогенов in vivo". Клетка. 49 (4): 465–75. Дои:10.1016/0092-8674(87)90449-1. PMID 3032456.
- ^ Siegel, P.M .; Шу, Вт; Кардифф, Р. Д .; Muller, W. J .; Массагуэ, Дж (2003). «Передача сигналов трансформирующего фактора роста бета нарушает индуцированный Neu онкогенез молочной железы, одновременно способствуя легочным метастазам». Труды Национальной академии наук. 100 (14): 8430–5. Дои:10.1073 / pnas.0932636100. ЧВК 166246. PMID 12808151.
- ^ Gunther, E.J .; Белка, Г.К .; Wertheim, G.B .; Ван, Дж; Hartman, J. L .; Боксер, Р. Б .; Ходош, Л. А. (2002). «Новая индуцируемая доксициклином система для трансгенного анализа биологии молочной железы». Журнал FASEB. 16 (3): 283–92. Дои:10.1096 / fj.01-0551com. PMID 11874978.
- ^ Подсыпанина, К; Politi, K; Беверли, Л. Дж .; Вармус, Х. Э. (2008). «Сотрудничество онкогенов в поддержании опухоли и рецидиве опухоли в опухолях молочной железы мышей, индуцированных Myc и мутантом Kras». Труды Национальной академии наук. 105 (13): 5242–7. Дои:10.1073 / pnas.0801197105. ЧВК 2278195. PMID 18356293.
- ^ Bierie, B; Stover, D.G .; Abel, T. W .; Chytil, A; Горска, А.Е .; Акре, М; Форрестер, Э; Ян, Л; Вагнер, К. У .; Моисей, Х. Л. (2008). «Трансформирующий фактор роста бета регулирует выживаемость клеток карциномы молочной железы и взаимодействие с прилегающим микроокружением». Исследования рака. 68 (6): 1809–19. Дои:10.1158 / 0008-5472.CAN-07-5597. PMID 18339861.
- ^ Коркмаз, Г; Lopes, R; Угальде, AP; Неведомская, Е; Han, R; Мячева, К; Zwart, W; Elkon, R; Агами, Р. (2016). Функциональная генетика проверяет элементы энхансера в геноме человека с помощью CRISPR-Cas9. Природа Биотехнологии 34, 192-198. DOI: 10.1038 / NBT.3450
- ^ Шринивас, S; Ватанабэ, Т; Lin, C. S .; William, C.M .; Танабэ, Й; Джесселл, Т. М .; Костантини, Ф (2001). «Репортерные штаммы Cre, полученные путем направленной вставки EYFP и ECFP в локус ROSA26». Биология развития BMC. 1: 4. Дои:10.1186 / 1471-213X-1-4. ЧВК 31338. PMID 11299042.
- ^ Liao, M. J .; Zhang, C.C .; Чжоу, B; Zimonjic, D. B .; Mani, S.A .; Каба, М; Гиффорд, А; Рейнхардт, Ф; Popescu, N.C .; Guo, W; Eaton, E.N .; Lodish, H. F .; Вайнберг, Р. А. (2007). «Обогащение популяции клеток молочной железы, которые образуют молочные железы и обладают репопуляционной активностью in vivo». Исследования рака. 67 (17): 8131–8. Дои:10.1158 / 0008-5472.CAN-06-4493. PMID 17804725.
- ^ Бисвас, S; Guix, M; Райнхарт, К; Dugger, T. C .; Chytil, A; Moses, H.L .; Freeman, M. L .; Артеага, К. Л. (2007). «Ингибирование TGF-бета с помощью нейтрализующих антител предотвращает индуцированное излучением ускорение метастатического прогрессирования рака». Журнал клинических исследований. 117 (5): 1305–13. Дои:10.1172/JCI30740. ЧВК 1838926. PMID 17415413.
- ^ Hüsemann, Y; Geigl, J. B.; Schubert, F; Musiani, P; Meyer, M; Burghart, E; Forni, G; Eils, R; Fehm, T; Riethmüller, G; Klein, C. A. (2008). "Systemic spread is an early step in breast cancer". Раковая клетка. 13 (1): 58–68. Дои:10.1016/j.ccr.2007.12.003. PMID 18167340.
- ^ Egeblad, M; Nakasone, E. S.; Werb, Z (2010). "Tumors as organs: Complex tissues that interface with the entire organism". Клетка развития. 18 (6): 884–901. Дои:10.1016/j.devcel.2010.05.012. ЧВК 2905377. PMID 20627072.
- ^ Entenberg, D; Wyckoff, J; Gligorijevic, B; Roussos, E. T.; Verkhusha, V. V.; Pollard, J. W.; Condeelis, J (2011). "Setup and use of a two-laser multiphoton microscope for multichannel intravital fluorescence imaging". Протоколы природы. 6 (10): 1500–20. Дои:10.1038/nprot.2011.376. ЧВК 4028841. PMID 21959234.
- ^ Alberini, Jean-Louis; Boisgard, Raphaël; Guillermet, Stéphanie; Siquier, Karine; Jego, Benoît; Thézé, Benoît; Urien, Saik; Rezaï, Keyvan; Menet, Emmanuelle (2016-08-01). "Multimodal In Vivo Imaging of Tumorigenesis and Response to Chemotherapy in a Transgenic Mouse Model of Mammary Cancer". Molecular Imaging and Biology. 18 (4): 617–626. Дои:10.1007/s11307-015-0916-7. ISSN 1860-2002. ЧВК 4927598. PMID 26630973.
внешняя ссылка
- http://www.la-press.com/tetracycline-regulated-systems-in-functional-oncogenomics-article-a200 A detailed overview of Tet-systems in functional oncogenomics