Гаплогруппа I-M253 - Haplogroup I-M253
Эта статья требует внимания специалиста по генетической истории человека. Конкретная проблема: Номенклатура гаплогрупп и субкладов. (Февраль 2017 г.) |
| Гаплогруппа I1 (M253) | |
|---|---|
| Возможное время происхождения | 3,170–4,600[1]-5,070 BP (сегодняшняя диверсификация)[2][3] (ранее 11000 л.[4] до 33000 лет назад[5])27 500 (диверсификация с I2-FGC77992)[1] |
| Возможное место происхождения | Северная Европа |
| Предок | Я* (M170) |
| Потомки | I1a (DF29 / S438); I1b (S249 / Z131); I1c (Y18119 / Z17925) |
| Определение мутаций | M253, M307.2 / P203.2, M450 / S109, P30, P40, L64, L75, L80, L81, L118, L121 / S62, L123, L124 / S64, L125 / S65, L157.1, L186, L187 |
Гаплогруппа I-M253, также известный как I1, это Y-хромосома гаплогруппа. Генетические маркеры, идентифицирующие I-M253, являются SNP M253, M307.2 / P203.2, M450 / S109, P30, P40, L64, L75, L80, L81, L118, L121 / S62, L123, L124 / S64, L125 / S65, L157.1, L186 и L187 .[6] Это основная ветвь Гаплогруппа I-M170 (Я*).
Считается, что гаплогруппа присутствовала среди европейских охотников-собирателей верхнего палеолита как второстепенная линия, но из-за ее отсутствия в образцах ДНК до неолита она не могла быть очень широко распространена. Образцы эпохи неолита I1 также очень редки, что предполагает быстрое рассеяние, связанное с эффектом основателя в Северный бронзовый век. Сегодня он достигает своих пиковых частот в Швеция (52 процента мужчин в Лен Вестра-Гёталанд ) и западный Финляндия (более 50 процентов в Сатакунта провинция).[7] В среднем по стране I-M253 встречается в 38-39% Шведский мужчины[8][9][10] 37% от норвежский язык мужчины[11][12] 32,8% от Датский мужчины[13] и около 28% Финский самцы.[14]
Гаплогруппа I-M253 является первичной ветвью гаплогруппы. Я* (I-M170), который присутствует в Европе с древних времен. Другая первичная ветвь I * - это I-M438, также известный как I2.
Все известные живые члены происходят от общего предка в 6 раз моложе общего предка с I2.[1]
До реклассификации в 2008 г.[15] группа была известна как I1a, имя, которое с тех пор было переназначено основной ветви, гаплогруппе I-DF29. Другими первичными ветвями I1 (M253) являются I1b (S249 / Z131) и I1c (Y18119 / Z17925).
Происхождение
Хотя гаплогруппа I1, скорее всего, расходилась с I * еще 27000 лет назад в Gravettian, до сих пор никакие ДНК-исследования не смогли обнаружить его у мезолитических охотников-собирателей. По состоянию на ноябрь 2020 года только 3 образца древней ДНК, относящиеся к более раннему периоду, чем нордический бронзовый век, были отнесены к гаплогруппе I1. Первый - это индивидуальный образец, обозначенный как BAB5, из Венгрии эпохи неолита.[16] Другой - человек, относящийся к среднему неолиту. Шассенская культура помечены как Cx161. Cx161 имел генетическое родство с другими современниками. Фермеры эпохи неолита Европы.[17] Кроме того, третий древний образец I1 взят от человека, найденного в курган захоронение, относящееся к эпохе позднего неолита в Скандинавии, обозначенное RISE179.[18] RISE179 имел генетическое родство с популяциями Культура шнуровой керамики и Культура Unetice.[19]
Несмотря на высокую частоту гаплогруппы I1 у современных скандинавов, I1 полностью отсутствует в образцах скандинавской мезолитической ДНК.[20][21] I1 впервые появляется в Скандинавии во время позднего неолита, но не увеличивается по частоте до начала века. Северный бронзовый век.[22][23][24][19]
Учитывая природу древних образцов ДНК Cx161 и RISE179, возможное объяснение того, почему гаплогруппа I1 до сих пор отсутствует среди мезолитических и неолитических охотников-собирателей в Скандинавии, заключается в том, что I1, возможно, присутствовал как второстепенная линия в центральноевропейских фермерских культурах неолита, таких как то Культура шаровидных амфор и Культура воронкообразных.[25] Позже он мог быть ассимилирован людьми культуры шнуровой керамики и привезен в Скандинавию как часть Культура боевого топора.[26][27][28]
Согласно исследованию, опубликованному в 2010 году, I-M253 возник между 3170 и 5000 лет назад в Энеолитическая Европа.[2] Новое исследование, проведенное в 2015 году, оценило его происхождение от 3 470 до 5070 лет назад или от 3 180 до 3760 лет назад с использованием двух разных методов.[3]
В 2007 году было высказано предположение, что первоначально он рассредоточился с территории, которая сейчас Дания.[13]Тем не мение, Проф. Д-р Кеннет Нордтведт, Государственный университет Монтаны по поводу MRCA в 2009 году написал в личном сообщении: «Мы не знаем, где существовал этот человек, но большая нижняя часть бассейна Эльбы кажется сердцем I1».
Последние результаты (сентябрь 2019 г.), опубликованные Y-полный предполагаю, что I1 (I-M253) был сформирован 27,500 лн (95 ДИ: 29.800 лн - 25.200 лн) с TMRCA 4.600 лн (95 CI: 5.200 лн - 4.000 лн).
Исследование 2014 года в Венгрии обнаружило останки двух человек из Линейная гончарная культура, один из которых, как было обнаружено, нес M253 SNP, который определяет гаплогруппу I1. Считается, что эта культура существовала от 7500 до 6500 лет назад.[29]
Структура
I-M253 (M253, M307.2 / P203.2, M450 / S109, P30, P40, L64, L75, L80, L81, L118, L121 / S62, L123, L124 / S64, L125 / S65, L157.1, L186 и L187) или же I1 [6]
- I-DF29 (DF29 / S438); I1a
- I-CTS6364 (CTS6364 / Z2336); I1a1
- FGC20030; I1a1a ~
- S4767; I1a1a1 ~
- I-M227; I1a1a1a1a
- A394; I1a1a2 ~
- Y11221; I1a1a3 ~
- A5338; I1a1a4 ~
- S4767; I1a1a1 ~
- CTS10028; I1a1b
- I-L22 (L22 / S142); I1a1b1
- CTS11651 / Z2338; I1a1b1a ~
- I-P109; I1a1b1a1
- I-Y3662; I1a1b1a1e ~
- I-S14887; I1a1b1a1e2 ~
- I-Y11203; I1a1b1a1e2d ~
- I-Y29630; I1a1b1a1e2d2 ~
- I-Y11203; I1a1b1a1e2d ~
- I-S14887; I1a1b1a1e2 ~
- I-Y3662; I1a1b1a1e ~
- CTS6017; I1a1b1a2
- I-L205 (L205.1 / L939.1 / S239.1); I1a1b1a3
- CTS6868; I1a1b1a4
- I-Z74; I1a1b1a4a
- CTS2208; I1a1b1a4a1 ~
- I-L287; I1a1b1a4a1a
- I-L258 (L258 / S335); I1a1b1a4a1a1
- I-L287; I1a1b1a4a1a
- I-L813; I1a1b1a4a2
- FGC12562; I1a1b1a4a3 ~
- CTS2208; I1a1b1a4a1 ~
- I-Z74; I1a1b1a4a
- I-P109; I1a1b1a1
- CTS11603 / S4744; I1a1b1b ~
- I-L300 (L300 / S241); I1a1b1b1a1
- CTS11651 / Z2338; I1a1b1a ~
- FGC10477 / Y13056; I1a1b2
- A8178, A8182, A8200, A8204; I1a1b3 ~
- F13534.2 / Y17263.2; I1a1b4 ~
- I-L22 (L22 / S142); I1a1b1
- FGC20030; I1a1a ~
- I-Z58 (S244 / Z58); I1a2
- I-Z59 (S246 / Z59); I1a2a
- I-Z60 (S337 / Z60, S439 / Z61, Z62); I1a2a1
- I-Z140 (Z140, Z141)
- I-L338
- I-F2642 (F2642)
- I-Z73
- I-L1302
- I-L573
- I-L803
- I-Z140 (Z140, Z141)
- I-Z382; I1a2a2
- I-Z60 (S337 / Z60, S439 / Z61, Z62); I1a2a1
- I-Z138 (S296 / Z138, Z139); I1a2b
- I-Z2541
- I-Z59 (S246 / Z59); I1a2a
- I-Z63 (S243 / Z63); I1a3
- I-BY151; I1a3a
- I-L849.2; I1a3a1
- I-BY351; I1a3a2
- I-CTS10345
- I-Y10994
- I-Y7075
- I-CTS10345
- I-S2078
- I-S2077
- I-Y2245 (Y2245 / PR683)
- I-L1237
- I-FGC9550
- I-S10360
- I-S15301
- I-Y7234
- I-L1237
- I-Y2245 (Y2245 / PR683)
- I-S2077
- I-BY62 (BY62); I1a3a3
- I-BY151; I1a3a
- I-CTS6364 (CTS6364 / Z2336); I1a1
- I-Z131 (Z131 / S249); I1b
- I-CTS6397; I1b1
- I-Z17943 (Y18119 / Z17925, S2304 / Z17937); I1c
Географическое распределение
I-M253 находится в самой высокой плотности в Северной Европе и других странах, которые испытали обширную миграцию из Северной Европы, либо в Период миграции, то Эпоха викингов, или современность. Он встречается во всех местах, захваченных норвежцами.
В современную эпоху значительные популяции I-M253 также укоренились в иммигрантских странах и бывших европейских колониях, таких как Соединенные Штаты, Австралия и Канада.
| численность населения | Размер образца | Я (всего) | I1 (I-M253) | I1a1a (I-M227) | Источник |
|---|---|---|---|---|---|
| Албанцы (Тирана ) | 55 | 21.82%=(12/55) | 3.6%=(2/55) | 0.0 | Battaglia et al. 2008 г. |
| Албанцы (Северная Македония ) | 64 | 17.2%=(11/64) | 4.7%=(3/64) | 0.0 | Battaglia et al. 2008 г. |
| Албанцы (Тирана ) Албанцы (Северная Македония ) | 55+64=119 | 19.33%=(23/119) | 4.2%=(5/119) | 0.0 | Battaglia et al. 2008 г. |
| Косовские албанцы (Приштина ) | 114 | 7.96%=(9/114) | 5.31%=(6/114) | 0.0 | Pericic et al. 2005 г. |
| Албанцы (Тирана ) Албанцы (Северная Македония ) Косовские албанцы (Приштина ) | 55+64+114=233 | 13.73%=(32/233) | 4.72%=(11/233) | 0.0 | Pericic et al. 2005 г. Battaglia et al. 2008 г. |
| Австрия | 43 | 9.3 | 2.3 | 0.0 | Андерхилл и др. 2007 г. |
| Беларусь: Витбск | 100 | 15 | 1.0 | 0.0 | Андерхилл и др. 2007 г. |
| Беларусь: Брест | 97 | 20.6 | 1.0 | 0.0 | Андерхилл и др. 2007 г. |
| Босния | 100 | 42 | 2.0 | 0.0 | Rootsi et al. 2004 г. |
| Болгария | 808 | 26.6 | 4.3 | 0.0 | Карачанак и др. 2013 |
| Чехия | 47 | 31.9 | 8.5 | 0.0 | Андерхилл и др. 2007 г. |
| Чехия | 53 | 17.0 | 1.9 | 0.0 | Rootsi et al. 2004 г. |
| Дания | 122 | 39.3% (48/122) | 32.8% (40/122) | 0.0 | Андерхилл и др. 2007 г. |
| Англия | 104 | 19.2 | 15.4 | 0.0 | Андерхилл и др. 2007 г. |
| Эстония | 210 | 18.6 | 14.8 | 0.5 | Rootsi et al. 2004 г. |
| Эстония | 118 | 11.9 | Lappalainen et al. 2008 г. | ||
| Финляндия (национальная) | 28.0 | Lappalainen et al. 2006 г. | |||
| Финляндия: Запад | 230 | 40% (92/230) | Lappalainen et al. 2008 г. | ||
| Финляндия: Восток | 306 | 19% (58/306) | Lappalainen et al. 2008 г. | ||
| Финляндия: регион Сатакунта | 50+ | Lappalainen et al. 20089 | |||
| Франция | 58 | 17.2 | 8.6 | 1.7 | Андерхилл и др. 2007 г. |
| Франция | 12 | 16.7 | 16.7 | 0.0 | Cann et al. 2002 г. |
| Франция (Нижняя Нормандия) | 42 | 21.4 | 11.9 | 0.0 | Rootsi et al. 2004 г. |
| Германия | 125 | 24 | 15.2 | 0.0 | Андерхилл и др. 2007 г. |
| Греция | 171 | 15.8 | 2.3 | 0.0 | Андерхилл и др. 2007 г. |
| Венгрия | 113 | 25.7 | 13.3 | 0.0 | Rootsi et al. 2004 г. |
| Ирландия | 100 | 11 | 6.0 | 0.0 | Андерхилл и др. 2007 г. |
| Казанские татары | 53 | 13.2 | 11.3 | 0.0 | Трофимова 2015 |
| Латвия | 113 | 3.5 | Lappalainen et al. 2008 г. | ||
| Литва | 164 | 4.9 | Lappalainen et al. 2008 г. | ||
| Нидерланды | 93 | 20.4 | 14 | 0.0 | Андерхилл и др. 2007 г. |
| Норвегия | 2826 | 31.5% (890/2826) | Евпедия 2017[нужен лучший источник ] | ||
| Россия (национальная) | 16 | 25 | 12.5 | 0.0 | Cann et al. 2002 г. |
| Россия: Псков | 130 | 16.9 | 5.4 | 0.0 | Андерхилл и др. 2007 г. |
| Россия: Кострома | 53 | 26.4 | 11.3 | 0.0 | Андерхилл и др. 2007 г. |
| Россия: Смоленск | 103 | 12.6 | 1.9 | 0.0 | Андерхилл и др. 2007 г. |
| Россия: Воронеж | 96 | 19.8 | 3.1 | 0.0 | Андерхилл и др. 2007 г. |
| Россия: Архангельск | 145 | 15.8 | 7.6 | 0.0 | Андерхилл и др. 2007 г. |
| Россия: казаки | 89 | 24.7 | 4.5 | 0.0 | Андерхилл и др. 2007 г. |
| Россия: карелы | 140 | 10 | 8.6 | 0.0 | Андерхилл и др. 2007 г. |
| Россия: карелы | 132 | 15.2 | Lappalainen et al. 2008 г. | ||
| Россия: Вепса | 39 | 5.1 | 2.6 | 0.0 | Андерхилл и др. 2007 г. |
| Словакия | 70 | 14.3 | 4.3 | 0.0 | Rootsi et al. 2004 г. |
| Словения | 95 | 26.3 | 7.4 | 0.0 | Андерхилл и др. 2007 г. |
| Швеция (национальная) | 160 | 35.6% (57/160) | Lappalainen et al. 2008 г. | ||
| Швеция (национальная) | 38.0 | Lappalainen et al. 2009 г. | |||
| Швеция: Вестра Гёталанд | 52 | Lappalainen et al. 2009 г. | |||
| Швейцария | 144 | 7.6 | 5.6 | 0.0 | Rootsi et al. 2004 г. |
| индюк | 523 | 5.4 | 1.1 | 0.0 | Андерхилл и др. 2007 г. |
| Украина: Львов | 101 | 23.8 | 4.9 | 0.0 | Андерхилл и др. 2007 г. |
| Украина: Иваново-Франков | 56 | 21.4 | 1.8 | 0.0 | Андерхилл и др. 2007 г. |
| Украина: Хмельниц | 176 | 26.2 | 6.1 | 0.0 | Андерхилл и др. 2007 г. |
| Украина: Черкассы | 114 | 28.1 | 4.3 | 0.0 | Андерхилл и др. 2007 г. |
| Украина: Белгород | 56 | 26.8 | 5.3 | 0.0 | Андерхилл и др. 2007 г. |
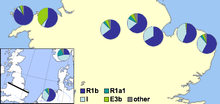
В 2002 году Майкл Уил и его коллеги опубликовали статью, в которой были показаны генетические доказательства популяционных различий между английским и валлийским населением, включая заметно более высокий уровень гаплогруппы I Y-ДНК в Англии, чем в Уэльсе. Они увидели в этом убедительное свидетельство массового вторжения англосаксонцев в восточную Великобританию из Северная Германия и Дания вовремя Период миграции.[30] Авторы предположили, что популяции с большой долей гаплогруппы I произошли из северной Германии или южной Скандинавии, особенно Дании, и что их предки мигрировали через Северное море с англосаксонскими миграциями и Датский Викинги. Основным утверждением исследователей было
что в то время требуется англо-саксонская иммиграция, которая затронет 50–100% мужского генофонда Центральной Англии. Тем не менее, мы отмечаем, что наши данные не позволяют нам отличить событие, которое просто добавляло мужской генофонд коренного населения Центральной Англии, от случая, когда коренные мужчины были перемещены в другие места, или случая, когда число самцов коренного населения сократилось ... Валлийская граница была скорее генетическим барьером для потока генов англосаксонской Y-хромосомы, чем Северное море ... Эти результаты показывают, что политическая граница может быть более важной, чем геофизическая, в генетическом структурировании популяции.

В 2003 году Кристиан Капелли и его коллеги опубликовали статью, в которой поддержали, но изменили выводы Уила и коллег.[31] В этой статье, в которой проводились выборки из Великобритании и Ирландии по сетке, было обнаружено меньшее различие между уэльским и английским выборками с постепенным уменьшением частоты гаплогруппы I с перемещением на запад в южной части Великобритании. Результаты показали авторам, что норвежские викинги-захватчики оказали сильное влияние на северную часть Британских островов, но что и английские, и континентальные шотландские образцы имеют немецкое / датское влияние.
Выдающиеся члены I-M253
Александр Гамильтон через генеалогию и тестирование его потомков (при условии, что фактическое отцовство соответствует его генеалогии), был помещен в гаплогруппу Y-ДНК I-M253.[32]
Биргер Ярл, «Герцог Швеции» из Восточно-геатишского дома Бьельбо, основатель Стокгольм; его останки были эксгумированы и исследованы в 2002 году и также оказались I-M253.[33]
Стинг был признан принадлежащим к гаплогруппе I1 сериалом PBS TV Finding Your Roots.[34]
Уильям Брэдфорд (губернатор) Mayflower, подтвержденный проектом ДНК Mayflower[35]
Уильям Брюстер (пассажир Mayflower) Mayflower, подтвержденный проектом ДНК Mayflower[35]
Маркеры

Ниже приведены технические характеристики известных мутаций SNP и STR гаплогруппы I-M253.
Название: M253[36]
- Тип: SNP
- Источник: M (Питер Андерхилл из Стэндфордский Университет )
- Позиция: ChrY: 13532101..13532101 (+ нить)
- Позиция (базовая пара): 283
- Общий размер (пары оснований): 400
- Длина: 1
- ISOGG HG: I1
- Праймер F (вперед 5 '→ 3'): GCAACAATGAGGGTTTTTTTG
- Праймер R (обратный 5 '→ 3'): CAGCTCCACCTCTATGCAGTTT
- YCC HG: I1
- Изменение (мутация) нуклеотидных аллелей: C к Т
Имя: M307[37]
- Тип: SNP
- Источник: M (Питер Андерхилл)
- Позиция: ChrY: 21160339..21160339 (+ нить)
- Длина: 1
- ISOGG HG: I1
- Праймер F: TTATTGGCATTTCAGGAAGTG
- Праймер R: GGGTGAGGCAGGAAAATAGC
- YCC HG: I1
- Изменение (мутация) нуклеотидных аллелей: грамм к А
Имя: P30[38]
- Тип: SNP
- Источник: PS (Майкл Хаммер из Университет Аризоны и Джеймс Ф. Уилсон, в Эдинбургском университете)
- Позиция: ChrY: 13006761..13006761 (+ нить)
- Длина: 1
- ISOGG HG: I1
- Праймер F: GGTGGGCTGTTTGAAAAAGA
- Праймер R: AGCCAAATACCAGTCGTCAC
- YCC HG: I1
- Изменение (мутация) нуклеотидных аллелей: с G на A
- Регион: ARSDP
Имя: P40[39]
- Тип: SNP
- Источник: PS (Майкл Хаммер и Джеймс Ф. Уилсон)
- Позиция: ChrY: 12994402..12994402 (+ нить)
- Длина: 1
- ISOGG HG: I1
- Праймер F: GGAGAAAAGGTGAGAAACC
- Праймер R: GGACAAGGGGCAGATT
- YCC HG: I1
- Изменение (мутация) нуклеотидных аллелей: C на T
- Регион: ARSDP
Смотрите также
Рекомендации
- ^ а б c https://yfull.com/tree/I1/
- ^ а б Педро Соарес, Алессандро Акилли, Орнелла Семино, Уильям Дэвис, Винсент Маколей, Ханс-Юрген Бандельт, Антонио Торрони и Мартин Б. Ричардс, Археогенетика Европы, Текущая биология, т. 20 (23 февраля 2010 г.), R174 – R183. yDNA гаплогруппа I: субклад I1, ДНК генеалогического древа,
- ^ а б Батини, Кьяра; Халласт, Пилле; Задик, Даниил; Дельсер, Пьерпаоло Майзано; Бенаццо, Андреа; Гиротто, Сильвия; Арройо-Пардо, Эдуардо; Cavalleri, Gianpiero L .; Де Книжфф, Питер; Дюпюи, Берит Мюре; Эриксен, Хайди А .; Король, Тури Э .; Де Мунайн, Адольфо Лопес; Лопес-Парра, Ана М .; Лутрадис, Афродита; Миласин, Елена; Новеллетто, Андреа; Памяв, Хорольма; Саджантила, Антти; Толун, Аслыхан; Винни, Брюс; Джоблинг, Марк А. (2015). «TMRCA основных гаплогрупп в Европе оценивается с использованием двух методов. Недавнее крупномасштабное расширение европейских родословных, показанное повторным упорядочиванием населения: Nature Communications: Nature Publishing Group». Nature Communications. 6: 7152. Дои:10.1038 / ncomms8152. ЧВК 4441248. PMID 25988751.
- ^ Рутси, Сиири; и другие. (2004). «Филогеография гаплогруппы I Y-хромосомы выявляет отдельные области доисторического потока генов в Европе» (PDF). Американский журнал генетики человека. 75 (1): 128–37. Дои:10.1086/422196. ЧВК 1181996. PMID 15162323. Архивировано из оригинал (PDF) на 2009-06-24. Получено 2008-03-20.
- ^ П.А. Андерхилл, Н.М. Майрес, С. Рутси, К.Т. Чоу, А.А. Лин, Р.П. Отиллар, Р.Кинг, Л.А. Животовский, О.Балановский, А.Пшеничнов, К.Х. Ричи, Л.Л. Кавалли-Сфорца, Т. Кивисилд, Р. Виллемс, С. Вудворд, Новые филогенетические отношения для гаплогруппы I Y-хромосомы: переоценка ее филогеографии и предыстории, в П. Мелларс, К. Бойл, О. Бар-Йосеф и К. Стрингер (ред.), Переосмысление эволюции человека (2007), стр. 33–42.
- ^ а б ISOGG, Y-ДНК Гаплогруппа I и ее субклады - 2017 (31 января 2017 г.).
- ^ Lappalainen, T .; Laitinen, V .; Salmela, E .; Andersen, P .; Huoponen, K .; Savontaus, M.-L .; Лахермо, П. (2008). «Миграционные волны в регион Балтийского моря». Анналы генетики человека. 72 (3): 337–48. Дои:10.1111 / j.1469-1809.2007.00429.x. PMID 18294359. S2CID 32079904.
- ^ Lappalainen, T .; Hannelius, U .; Salmela, E .; von Döbeln, U .; Lindgren, C.M .; Huoponen, K .; Savontaus, M.-L .; Kere, J .; Лахермо, П. (2009). "Структура населения в современной Швеции: анализ Y-хромосомы и митохондрий ДНК". Анналы генетики человека. 73 (1): 61–73. Дои:10.1111 / j.1469-1809.2008.00487.x. PMID 19040656. S2CID 205598345.
- ^ https://www.familytreedna.com/public/Sweden?iframe=ymap
- ^ Lappalainen, T .; Laitinen, V .; Salmela, E .; Andersen, P .; Huoponen, K .; Savontaus, M.-L .; Лахермо, П. (май 2008 г.). «Миграционные волны в регион Балтийского моря». Анналы генетики человека. 72 (Pt 3): 337–348. Дои:10.1111 / j.1469-1809.2007.00429.x. ISSN 0003-4800. PMID 18294359.
- ^ Дюпюи, Берит Мюре; Стенерсен, Маргурет; Лу, Тим Т .; Олайсен, Бьёрнар (01.12.2006). «Географическая неоднородность линий Y-хромосомы в Норвегии». Международная криминалистическая экспертиза. 164 (1): 10–19. Дои:10.1016 / j.forsciint.2005.11.009. ISSN 0379-0738. PMID 16337760.
- ^ "FamilyTreeDNA - Проект ДНК Норвегии - Norgesprosjektet". www.familytreedna.com. Получено 2020-11-26.
- ^ а б Питер А. Андерхилл и др., Новые филогенетические отношения для гаплогруппы I Y-хромосомы: переоценка ее филогеографии и предыстории, в Переосмысление человеческой революции (2007), стр. 33–42. П. Мелларс, К. Бойл, О. Бар-Йосеф, К. Стрингер (ред.) Институт археологических исследований Макдональда, Кембридж, Великобритания.
- ^ Лаппалайнен Т., Койвумяки С., Салмела Э., Хуопонен К., Систонен П., Савонтаус М.Л., Лахермо П .; 2006, "Региональные различия между финнами: перспектива Y-хромосомы", Ген т. 376, нет. 2. С. 207–15.
- ^ Карафет, Татьяна М .; Mendez, F. L .; Meilerman, M. B .; Андерхилл, П. А .; Zegura, S.L .; Хаммер, М. Ф. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека». Геномные исследования. 18 (5): 830–38. Дои:10.1101 / gr.7172008. ЧВК 2336805. PMID 18385274.
- ^ Сеченьи-Надь, Анна; Брандт, Гвидо; Кирл, Виктория; Якуч, Янош; Хаак, Вольфганг; Мёллер-Рикер, Сабина; Келер, Китти; Менде, Балаж; Фехер, Марк; Оросс, Кристиан; Палуч, Тибор; Osztás, Anett; Поцелуй, Виктория; Палфи, Дьёрдь; Мольнар, Эрика; Себек, Каталин; Чене, Андраш; Палуч, Тибор; Шлаус, Марио; Новак, Марио; Печина-Шлаус, Нивес; Ősz, Бригитта; Войсек, Ванда; Somogyi, Krisztina; Тот, Габор; Кромер, Бернд; Банффи, Эстер; Альт, Курт (2014). «Отслеживание генетического происхождения первых фермеров Европы позволяет лучше понять их социальную организацию». Дои:10.1101/008664. S2CID 196648568. Цитировать журнал требует
| журнал =(помощь) - ^ Брунель, Саманта; Беннет, Э. Эндрю; Карден, Лоран; Гарро, Дэмиен; Эмам, Элен Барран; Бейлиер, Александр; Булестен, Бруно; Ченал, Фанни; Чесельски, Эльза; Конвертини, Фабьен; Дедет, Бернард (2020-06-09). «Древние геномы современной Франции раскрывают 7000 лет ее демографической истории». Труды Национальной академии наук. 117 (23): 12791–12798. Дои:10.1073 / pnas.1918034117. ISSN 0027-8424. ЧВК 7293694. PMID 32457149.
- ^ Аллентофт, Мортен Э .; Сикора, Мартин; Сьегрен, Карл-Йоран; Расмуссен, Саймон; Расмуссен, Мортен; Стендеруп, Джеспер; Дамгаард, Питер Б .; Шредер, Ханнес; Альстрём, Торбьёрн; Виннер, Лассе; Маласпинас, Анна-Сапфо; Маргарян, Ашот; Хайэм, Том; Чивалл, Дэвид; Линнеруп, Нильс; Харвиг, Лиза; Барон, Юстина; Casa, Philippe Della; Домбровски, Павел; Даффи, Пол Р .; Эбель, Александр В .; Епимахов Андрей; Фрей, Карин; Фурманек, Мирослав; Гралак, Томаш; Громов Андрей; Гронкевич, Станислав; Grupe, Gisela; Хайду, Тамаш; и другие. (2015). «Популяционная геномика Евразии бронзового века». Природа. 522 (7555): 167–172. Bibcode:2015Натура.522..167A. Дои:10.1038 / природа14507. PMID 26062507. S2CID 4399103.
- ^ а б Аллентофт, Мортен Э .; Сикора, Мартин; Сьегрен, Карл-Йоран; Расмуссен, Саймон; Расмуссен, Мортен; Стендеруп, Джеспер; Дамгаард, Питер Б .; Шредер, Ханнес; Альстрём, Торбьёрн; Виннер, Лассе; Маласпинас, Анна-Сапфо (июнь 2015 г.). «Популяционная геномика Евразии бронзового века». Природа. 522 (7555): 167–172. Bibcode:2015Натура.522..167A. Дои:10.1038 / природа14507. ISSN 1476-4687. PMID 26062507. S2CID 4399103.
- ^ Günther, T .; Malmström, H .; Svensson, E.M .; Омрак, А .; Sánchez-Quinto, F .; Килинч, Г. М .; Krzewińska, M .; Eriksson, G .; Fraser, M .; Edlund, H .; Munters, A.R .; Coutinho, A .; Simões, L.G .; Vicente, M .; Sjölander, A .; Янсен Селлевольд, Б .; Jørgensen, R .; Claes, P .; Shriver, M.D .; Valdiosera, C .; Netea, M. G .; Apel, J .; Lidén, K .; Скар, Б .; Storå, J .; Götherström, A .; Якобссон, М. (2018). «Популяционная геномика мезолитической Скандинавии: изучение путей ранней послеледниковой миграции и адаптации к высоким широтам». PLOS Биология. 16 (1): e2003703. Дои:10.1371 / journal.pbio.2003703. ЧВК 5760011. PMID 29315301.
- ^ Malmström, H .; Günther, T .; Svensson, E.M .; Юрас, А .; Fraser, M .; Munters, A.R .; Тырв, М .; Lindström, J .; Götherström, A .; Storå, J .; Якобссон, М .; Якобссон, Маттиас (2019). «Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широким кругозором шнуровой керамики». Ход работы. Биологические науки. 286 (1912). Дои:10.1098 / rspb.2019.1528. ЧВК 6790770. PMID 31594508.
- ^ Санчес-Кинто, Федерико; Мальмстрем, Хелена; Фрейзер, Магдалена; Гирдланд-Флинк, Линус; Свенссон, Эмма М .; Simões, Luciana G .; Джордж, Роберт; Холлфельдер, Нина; Буренхульт, Горан; Благородный, Гордон; Бриттон, Кейт (2019-05-07). «Мегалитические гробницы в западной и северной неолитической Европе были связаны с родственным обществом». Труды Национальной академии наук. 116 (19): 9469–9474. Дои:10.1073 / pnas.1818037116. ISSN 0027-8424. ЧВК 6511028. PMID 30988179.
- ^ Скоглунд, Понт; Мальмстрем, Хелена; Омрак, Айча; Рагхаван, Маанаса; Вальдиосера, Кристина; Гюнтер, Торстен; Холл, Пер; Тамбец, Кристина; Парик, Юри; Сьегрен, Карл-Йоран; Апель, янв (16.05.2014). «Геномное разнообразие и различия в составе примесей у скандинавских собирателей и фермеров каменного века». Наука. 344 (6185): 747–750. Bibcode:2014Наука ... 344..747С. Дои:10.1126 / science.1253448. ISSN 0036-8075. PMID 24762536. S2CID 206556994.
- ^ Карлссон, Андреас О .; Валлерстрём, Томас; Гетерстрём, Андерс; Холмлунд, Гунилла (август 2006 г.). «Разнообразие Y-хромосомы в Швеции - долгосрочная перспектива». Европейский журнал генетики человека. 14 (8): 963–970. Дои:10.1038 / sj.ejhg.5201651. ISSN 1476-5438. PMID 16724001. S2CID 23227271.
- ^ https://www.eupedia.com/europe/Haplogroup_I1_Y-DNA.shtml#nordic
- ^ Мальмстрем, Хелена; Гилберт, М. Томас П .; Thomas, Mark G .; Брандстрём, Микаэль; Сторо, Ян; Мольнар, Петра; Андерсен, Пернилле К .; Бендиксен, Кристиан; Холмлунд, Гунилла; Гетерстрём, Андерс; Виллерслев, Эске (3 ноября 2009 г.). «Древняя ДНК показывает отсутствие преемственности между неолитическими охотниками-собирателями и современными скандинавами». Текущая биология. 19 (20): 1758–1762. Дои:10.1016 / j.cub.2009.09.017. ISSN 0960-9822. PMID 19781941. S2CID 9487217.
- ^ «Первые фермеры Скандинавии обменялись терминологией с индоевропейцами». Phys.org. Получено 2020-11-23.
- ^ Мальмстрем, Хелена; Гюнтер, Торстен; Свенссон, Эмма М .; Юрас, Анна; Фрейзер, Магдалена; Munters, Arielle R .; Поспешный, Лукаш; Тырв, Мари; Линдстрем, Джонатан; Гетерстрём, Андерс; Сторо, Ян; Якобссон, Маттиас (2019). «Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широким кругозором шнуровой керамики». Труды Королевского общества B: биологические науки. 286 (1912). Дои:10.1098 / rspb.2019.1528. ЧВК 6790770. PMID 31594508.
- ^ «Отслеживание генетического происхождения первых фермеров Европы позволяет лучше понять их социальную организацию». bioRxiv 10.1101/008664.
- ^ Уил, Майкл Э .; Weiss, Deborah A .; Jager, Rolf F .; Брэдман, Нил; Томас, Марк Г. (2002). "Свидетельства об англосаксонской массовой миграции по Y-хромосоме". Молекулярная биология и эволюция. 19 (7): 1008–21. Дои:10.1093 / oxfordjournals.molbev.a004160. PMID 12082121.
- ^ Капелли, Кристиан; Рыжая, Никола; Абернети, Джулия К .; Гратрикс, Фиона; Уилсон, Джеймс Ф .; Моэн, Торольф; Хервиг, Тор; Ричардс, Мартин; Штумпф, Майкл П.Х .; и другие. (2003). "Перепись Y-хромосомы на Британских островах" (PDF). Текущая биология. 13 (11): 979–84. Дои:10.1016 / S0960-9822 (03) 00373-7. PMID 12781138. S2CID 526263.
- ^ "ДНК отца-основателя". isogg.org.
- ^ Мальмстрем, Хелена; Вретемарк, Мария; Тильмар, Андреас; Дурлинг, Микаэль Брандстрём; Скоглунд, Понт; Гилберт, М. Томас П .; Виллерслев, Эске; Холмлунд, Гунилла; Гетерстрём, Андерс (20 января 2012 г.). «В поисках основателя Стокгольма - исследование родства на основе Y-хромосомной, аутосомной и митохондриальной ДНК». Анналы анатомии - Anatomischer Anzeiger. Спецвыпуск: Древняя ДНК. 194 (1): 138–145. Дои:10.1016 / j.aanat.2011.03.014. ISSN 0940-9602. PMID 21596538.
- ^ Британское вторжение В поисках корней
- ^ а б "Проект ДНК Мэйфлауэр". mayflowerdna.org. Получено 2020-11-23.
- ^ snpdev. "Отчет кластера эталонного SNP (refSNP): rs9341296". nih.gov.
- ^ snpdev. «Отчет кластера эталонного SNP (refSNP): rs13447354». nih.gov.
- ^ P30[постоянная мертвая ссылка ]
- ^ P40[постоянная мертвая ссылка ]
Источники
- Вильяльба-Муко, Ванесса; и другие. (1 апреля 2019 г.). "Выживание предков позднего плейстоцена охотников-собирателей на Пиренейском полуострове". Текущая биология. Cell Press. 29 (2): 1169–117. Дои:10.1016 / j.cub.2019.02.006. PMID 30880015. S2CID 76663708. Получено 12 ноября, 2020.
- Аллентофт и др. (Июнь 2015 г.) Популяционная геномика Евразии бронзового века https://www.researchgate.net/publication/278327861_Population_genomics_of_Bronze_Age_Eurasia
- Брюнель и др. (Июнь 2020 г.) Древние геномы современной Франции раскрывают 7000-летнюю демографическую историю https://www.pnas.org/content/117/23/12791
внешняя ссылка
- Базы данных гаплогруппы I
- Проект гаплогруппы I1 в FTDNA
- Датский региональный проект ДНК Демеса в FTDNA
- Гаплогруппа I-P109 Project
- Проект ДНК Британских островов
- Общие базы данных Y-ДНК
Существует несколько баз данных открытого доступа с использованием I-M253, в том числе:
- http://www.eupedia.com/europe/european_y-dna_haplogroups.shtml
- http://www.semargl.me/[постоянная мертвая ссылка ]
- http://www.ysearch.org/
- http://www.yhrd.org/
- http://www.yfull.com/tree/I1/
| Филогенетическое древо человека Гаплогруппы ДНК Y-хромосомы [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-хромосомный Адам " | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| грамм | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | ЧАС | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| я | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | Т | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | п [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| НЕТ | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | О | Q | р | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||