Гаструляция - Gastrulation
| Гаструляция | |
|---|---|
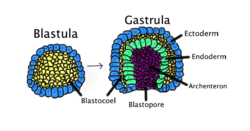 Гаструляция происходит, когда бластула, состоящая из одного слоя, загибается внутрь и увеличивается, образуя гаструлу. Эта диаграмма имеет цветовую маркировку: эктодерма - синий; энтодерма зеленая; бластоцель (желточный мешок) желтый; и архентерон (кишечник), пурпурный. | |
| Идентификаторы | |
| MeSH | D054262 |
| Анатомическая терминология | |
В биология развития, гаструляция это этап на ранней стадии эмбриональное развитие из большинства животные, во время которого бластула (однослойная полая сфера из клетки ) реорганизуется в многослойную структуру, известную как гаструла. Перед гаструляцией эмбрион является непрерывным эпителиальный лист ячеек; к концу гаструляции зародыш начался дифференциация установить четкие клеточные линии, установите основные оси тела (например, дорсально-вентральный, передне-задний ) и интернализовали один или несколько типов клеток, включая предполагаемый кишка.
В триплобластический организмов гаструла трехслойная («трехслойная»). Эти три ростковые отростки известны как эктодерма (Наружный слой), мезодерма (средний слой) и энтодерма (внутренний слой).[1][2] В диплобластический организмы, такие как Книдария и Гребневик, гаструла имеет только эктодерму и энтодерму. Два слоя также иногда называют гипобласт и эпибласт.[3]
Гаструляция происходит после расщепление и формирование бластула. После гаструляции следует органогенез, когда отдельные органы развиваются внутри новообразованных зародышевых листков.[4] Каждый слой порождает определенные ткани и органы в развивающемся эмбрионе.
- Эктодерма дает начало эпидермис, то нервная система, и к нервный гребень у позвоночных.
- Энтодерма дает начало эпителий из пищеварительная система и дыхательная система, и органы, связанные с пищеварительная система, такой как печень и поджелудочная железа.
- Мезодерма дает начало множеству типов клеток, таких как мышца, кость, и соединительная ткань. У позвоночных производные мезодермы включают нотохорда, то сердце, кровь и кровеносный сосуд, то хрящ из ребра и позвонки, а дерма.[5]
После гаструляции клетки в организме либо организуются в слои связанных клеток (как в эпителий ), или в виде сетки изолированных ячеек, например мезенхима.[2][6]
Хотя паттерны гаструляции сильно различаются в животном мире, их объединяет пять основных типов клеточных движений[7] которые возникают во время гаструляции:
Термины «гаструла» и «гаструляция» были изобретены Эрнст Геккель, в его работе 1872 г. «Биология известковых губок».[8] Льюис Вулперт, первопроходец в области биологии развития в этой области, был отмечен тем, что заметил, что «не рождение, брак или смерть, а гаструляция действительно является самым важным временем в вашей жизни».[9]

Классические модельные системы
Гаструляция сильно различается в животном мире, но имеет общие черты. Гаструляция изучалась на многих животных, но некоторые модели использовались дольше, чем другие. Кроме того, легче изучать развитие животных, которые развиваются вне матери. К животным, гаструляция которых изучена наиболее подробно, относятся:
Протостомы против дейтеростомов
В различие между протостомы и дейтеростомы основан на направлении развития рта (стомы) по отношению к бластопор. Протостом происходит от греческого слова protostoma, означающего «первый рот» (πρώτος + στόμα), тогда как этимология Второстома - «второй рот» от слов второй и рот (δεύτερος + στόμα).
Основные различия между дейтеростомами и протостомами обнаруживаются в эмбриональном развитии:
- Рот / анус
- Расщепление
- Протостомы имеют то, что известно как спиральный разрез который определенный, что означает, что судьба клеток определяется по мере их образования.
- Второстомы имеют то, что известно как радиальный разрез то есть неопределенный.
Морские ежи
Морские ежи Euechinoidea были важной модельной системой в биологии развития с 19 века.[10] Их гаструляция часто считается архетипом дейтеростомов беспозвоночных.[11] Эксперименты наряду с компьютерным моделированием были использованы для получения знаний о гаструляции морского ежа. Недавнее моделирование показало, что плоской клеточной полярности достаточно, чтобы управлять гаструляцией морского ежа.[12]
Определение зародышевого слоя
Морские ежи демонстрируют очень стереотипные модели расщепления и клеточные судьбы. На хранение матери мРНК установить организующий центр эмбриона морского ежа. Канонический Wnt и Delta-Notch сигнализация прогрессивно разделяет прогрессивную энтодерму и мезодерму.[13]
Интернализация клеток
В морские ежи первые клетки, которые усвоятся, являются первичными мезенхима ячейки (ЧВК), которые имеют скелетогенный судьба, попадающая на стадии бластулы. Гаструляция - интернализация будущего энтодерма и не скелетогенный мезодерма - начинается вскоре после этого с инвагинации и других перестроек клеток вегетативного полюса, которые вносят примерно 30% в окончательный архентерон длина. В окончательная длина кишки зависит от перестройки клеток внутри архентерона.[14]
Амфибии
В лягушка, Xenopus использовался как модельный организм для изучения гаструляции.
Нарушение симметрии
Сперма вносит один из двух митотические астры необходимо для завершения первого расщепления. Сперма может попасть в любую половину яйцеклетки животного, но ее точная точка входа нарушит радиальную симметрию яйцеклетки за счет организации цитоскелет. Перед первым делением кора яйца вращается относительно внутреннего цитоплазма согласованными действиями микротрубочки, в процессе, известном как кортикальное вращение. Это смещение приводит в контакт материнские детерминанты судьбы клеток из экваториальной цитоплазмы и вегетативной коры, и вместе эти детерминанты устанавливают организатор. Таким образом, органайзером станет участок на вегетативной стороне напротив точки входа сперматозоидов.[15] Хильде Мангольд, работая в лаборатории Ганс Спеманн, продемонстрировали, что этот особый «организатор» эмбриона необходим и достаточен для индукции гаструляции.[16][17][18]
Определение зародышевого слоя
Спецификация энтодермы зависит от перестройки материнских детерминант, что приводит к нуклеаризации Бета-катенин. Мезодерма индуцированный путем передачи сигналов от презумптивной энтодермы к клеткам, которые в противном случае стали бы эктодермой.[15]
Интернализация клеток
В спинной губа бластопора является механической движущей силой гаструляции. Первые признаки инвагинации это видео гаструляции лягушки это задняя губа.
Передача сигналов клетки
В лягушке, Ксеноп, один из сигналов - ретиноевая кислота (RA).[19] Передача сигналов RA в этом организме может влиять на формирование энтодермы и, в зависимости от времени передачи сигналов, может определять судьбу, будь то панкреатическая, кишечная или респираторная. Другие сигналы, такие как Wnt и BMP, также играют роль в респираторной судьбе Xenopus путем активации индикаторов клеточного происхождения.[19]
Амниот
Обзор
В амниот (рептилии, птицы и млекопитающие), гаструляция включает создание бластопора, входа в архентерон. Обратите внимание, что бластопор не является отверстием в бластоцель, пространство внутри бластула, но представляет собой новое углубление, которое сближает существующие поверхности бластулы. В амниот, гаструляция происходит в следующей последовательности: (1) эмбрион становится асимметричный; (2) примитивная полоса формы; (3) клетки из эпибласт на примитивная полоса пройти эпителиально-мезенхимальный переход и входить на примитивная полоса сформировать ростковые отростки.[5]
Нарушение симметрии
При подготовке к гаструляции эмбрион должен стать асимметричным по обеим сторонам. проксимально-дистальная ось и передне-задняя ось. Проксимально-дистальная ось образуется, когда клетки эмбриона образуют «яйцевой цилиндр», который состоит из внеэмбриональных тканей, которые дают начало структурам, подобным плацента, на проксимальном конце и эпибласт на дистальном конце. Многие сигнальные пути способствуют этой реорганизации, в том числе BMP, FGF, узловой, и Wnt. Висцеральная энтодерма окружает эпибласт. В дистальный висцеральная энтодерма (DVE) мигрирует в передний часть эмбриона, образующая «переднюю висцеральную энтодерму» (AVE). Это нарушает передне-заднюю симметрию и регулируется узловой сигнализация.[5]
Определение зародышевого слоя
В примитивная полоса формируется в начале гаструляции и находится на стыке между внеэмбриональной тканью и эпибласт на задней стороне эмбриона и на месте вторжение.[20] Формирование примитивная полоса зависит от узловой сигнализация[5] в Серп Коллера внутри клеток, способствующих примитивной полосе и BMP4 передача сигналов из экстраэмбриональной ткани.[20][21] Более того, Cer1 и Левша1 ограничить примитивную полосу подходящим местом, противодействуя узловой сигнализация.[22] Регион, определяемый как примитивная полоса продолжает расти к дистальному концу.[5]
На ранних стадиях развития примитивная полоса - это структура, которая установит двусторонняя симметрия, определить место гаструляции и инициировать формирование зародышевого листка. Чтобы сформировать полосу, рептилии, птицы и млекопитающие размещают мезенхимные клетки вдоль предполагаемой средней линии, устанавливая первую ось эмбриона, а также место, куда клетки будут проникать и мигрировать в процессе гаструляции и формирования зародышевого листка.[23] Примитивная полоса проходит через эту среднюю линию и образует переднезаднюю ось тела,[24] став первым событием нарушения симметрии в эмбрион, и знаменует начало гаструляции.[25] Этот процесс включает проникновение предшественников мезодермы и энтодермы и их миграцию в их окончательное положение,[24][26] где они будут дифференцироваться на три зародышевых листка.[23] Локализация клеточной адгезии и сигнальной молекулы бета-катенин критически важен для правильного образования области организатора, ответственной за инициирование гаструляции.
Интернализация клеток
Чтобы клетки переместились из эпителий из эпибласт сквозь примитивная полоса чтобы сформировать новый слой, клетки должны пройти эпителиально-мезенхимальный переход (EMT), чтобы потерять свои эпителиальные характеристики, такие как межклеточная адгезия. FGF сигнализация необходима для правильного EMT. FGFR1 необходим для повышения SNAI1, который регулирует вниз E-кадгерин, вызывая потерю клеточной адгезии. После ЕМТ клетки входить сквозь примитивная полоса и распространиться, чтобы сформировать новый слой ячеек или присоединиться к существующим слоям. FGF8 вовлечен в процесс этого рассеивания от примитивная полоса.[22]
Передача сигналов клетки
Есть определенные сигналы, которые играют роль в детерминации и формировании трех зародышевых листков, таких как FGF, RA и Wnt.[19] У млекопитающих, таких как мыши, передача сигналов RA может играть роль в формировании легких. Если RA недостаточно, в производстве легких будет ошибка. RA также регулирует респираторную компетентность в этой модели мышей.
Передача клеточных сигналов, управляющая гаструляцией
Во время гаструляции клетки дифференцируются в эктодерму или мезендодерму, которая затем разделяется на мезодерму и энтодерму.[19] Энтодерма и мезодерма формируются за счет узловой передачи сигналов. Узловая передача сигналов использует лиганды, которые являются частью семейства TGFβ. Эти лиганды будут сигнализировать о трансмембранных рецепторах серин / треонинкиназы, которые затем будут фосфорилировать Smad2 и Smad3. Затем этот белок присоединяется к Smad4 и перемещается в ядро, где начинают транскрибироваться гены мезэндодермы. Путь Wnt вместе с β-catenin играет ключевую роль в передаче сигналов узлов и формировании энтодермы.[27] Факторы роста фибробластов (FGF), канонический путь Wnt, костный морфогенетический белок (BMP) и ретиноевая кислота (RA) - все они важны для формирования и развития энтодермы.[19] FGF важны в производстве гена гомеобокса, который регулирует раннее анатомическое развитие. Передача сигналов BMP играет роль в печени и способствует ее судьбе. Передача сигналов RA также индуцирует гены гомеобокса, такие как Hoxb1 и Hoxa5. У мышей, если отсутствует передача сигналов RA, у мыши не разовьются легкие.[19] Передача сигналов RA также имеет множество применений при формировании органов глоточных дуг, передней и задней кишки.[19]
Гаструляция in vitro
Было предпринято несколько попыток понять процессы гаструляции, используя in vitro методы параллельно и дополняющие исследования на эмбрионах, обычно с использованием 2D[28][29][30] и 3D ячейка (Эмбриональные органоиды ) методы культивирования[31][32][33][34] с помощью Эмбриональные стволовые клетки (ESC) или индуцированные плюрипотентные стволовые клетки (ИПСК). Это связано с рядом явных преимуществ при использовании протоколов на основе тканевых культур, некоторые из которых включают снижение стоимости связанных in vivo работа (тем самым сокращая, заменяя и улучшая использование животных в экспериментах; 3R ), способность точно применять агонисты / антагонисты пространственно и временно определенным образом[32][33] что может быть технически сложно выполнить во время гаструляции. Однако для контекста важно связать наблюдения в культуре с процессами, происходящими в эмбрионе.
Чтобы проиллюстрировать это, управляемая дифференциация ESC мыши привела к созданию примитивная полоса -подобные клетки, которые демонстрируют многие характеристики клеток эпибласта, которые проходят через примитивную полосу[28] (например, временный брахьюри регуляции и клеточных изменений, связанных с эпителиально-мезенхимальный переход[28]), и человеческие ЭСК, культивируемые на микропланах, обработанные BMP4, может генерировать шаблон пространственной дифференциации, аналогичный расположению ростковые отростки в человеческом эмбрионе.[29][30] Наконец, используя 3D эмбриоидное тело - и органоид -основанные методы, небольшие агрегаты ЭСК мыши (Эмбриональные органоиды или гаструлоиды ) способны показать ряд процессов раннего развития эмбриона млекопитающих, таких как нарушение симметрии, поляризация экспрессии генов, движения, подобные гаструляции, удлинение оси и образование всех трех осей эмбриона (переднезадней, дорсовентральной и левой-правой осей) .[31][32][33][35]
Смотрите также
Рекомендации
Примечания
- ^ Mundlos 2009: п. 422
- ^ а б Макгиди, 2004: с. 34
- ^ Джонатон М.В., Slack (2013). Основная биология развития. Западный Сассекс, Великобритания: Wiley-Blackwell. п. 122. ISBN 978-0-470-92351-1.
- ^ Холл, 1998: стр.132-134
- ^ а б c d е Арнольд и Робинсон, 2009 г.
- ^ Холл, 1998: п. 177
- ^ Гилберт 2010: стр. 164.
- ^ Ересковский 2010: п. 236
- ^ Вольперт Л (2008) Торжество эмбриона. Courier Corporation, стр. 12. ISBN 9780486469294
- ^ Лаубихлер, доктор медицины и Дэвидсон, Э. Х. (2008). «Длительный эксперимент Бовери: мерогоны морского ежа и установление роли ядерных хромосом в развитии». «Биология развития». «314 (1): 1-11». "DOI: 10.1016 / j.ydbio.2007.11.024".
- ^ Макклей, Дэвид Р .; Gross, J.M .; Диапазон, Райан; Peterson, R.E .; Брэдхэм, Синтия (2004). «Глава 9: Гаструляция морского ежа». В Стерне, Клаудио Д. (ред.). Гаструляция: от клеток к эмбрионам. Лабораторный пресс Колд-Спринг-Харбор. С. 123–137. ISBN 978-0-87969-707-5.
- ^ Нильсен, Бьярке Фрост; Ниссен, Сайлас Бойе; Снеппен, Ким; Матизен, Иоахим; Трусина, Ала (21 февраля 2020 г.). «Модель, связывающая форму и полярность клеток с органогенезом». iScience. 23 (2): 100830. Bibcode:2020iSci ... 23j0830N. Дои:10.1016 / j.isci.2020.100830. ЧВК 6994644. PMID 31986479. S2CID 210934521.
- ^ Макклей, Д. Р. 2009. Расщепление и гаструляция у морского ежа. eLS. Дои:10.1002 / 9780470015902.a0001073.pub2
- ^ Хардин Дж. Д. (1990). «Контекстно-зависимое поведение клеток во время гаструляции» (PDF). Семин. Dev. Биол. 1: 335–345.
- ^ а б Гилберт, Скотт Ф. (2000). "Формирование оси у земноводных: феномен организатора, прогрессивное определение осей земноводных". Биология развития. Sinauer Associates.
- ^ Гилберт, Скотт Ф. (2000). «Рисунок 10.20, [Организация вторичной оси ...]». www.ncbi.nlm.nih.gov. Получено 1 июня 2020.
- ^ Спеманн Х., Мангольд Х. (1924). "Über Induktion von Embryonanlagen durch Implantation artfremder Organisatoren". Ру Арка. F. Entw. Мех. 100 (3–4): 599–638. Дои:10.1007 / bf02108133. S2CID 12605303.
- ^ Де Роберти Эдвард (2006). «Организатор Спеманна и саморегуляция у эмбрионов амфибий». Обзоры природы Молекулярная клеточная биология. 7 (4): 296–302. Дои:10.1038 / nrm1855. ЧВК 2464568. PMID 16482093.
- ^ а б c d е ж грамм Зорн А., Уэллс Дж. (2009). «Развитие эндодермы позвоночных и формирование органов». Анну Рев Селл Дев Биол. 25: 221–251. Дои:10.1146 / annurev.cellbio.042308.113344. ЧВК 2861293. PMID 19575677.
- ^ а б Там и Берингер, 1997 г.
- ^ Катала, 2005 г .: п. 1535
- ^ а б Tam, P.P .; Лёбель, Д.А. (2007). «Функция генов в эмбриогенезе мышей: приготовьтесь к гаструляции». Нат Рев Жене. 8 (5): 368–81. Дои:10.1038 / nrg2084. PMID 17387317. S2CID 138874.
- ^ а б Mikawa T, Poh AM, Kelly KA, Ishii Y, Reese DE (2004). «Индукция и формирование первичной полоски, организующего центра гаструляции в амниоте». Дев Дин. 229 (3): 422–32. Дои:10.1002 / dvdy.10458. PMID 14991697. S2CID 758473.
- ^ а б Даунс КМ. (2009). «Загадочная примитивная полоса: преобладающие представления и проблемы, касающиеся оси тела млекопитающих». BioEssays. 31 (8): 892–902. Дои:10.1002 / bies.200900038. ЧВК 2949267. PMID 19609969.
- ^ Чуай М., Цзэн В., Ян X, Бойченко В., Стекольщик Дж. А., Вейер С. Дж. (2006). «Движение клеток при формировании примитивной полосы у цыплят». Dev. Биол. 296 (1): 137–49. Дои:10.1016 / j.ydbio.2006.04.451. ЧВК 2556955. PMID 16725136.
- ^ Чуай М., Вейджер CJ (2008). Механизмы формирования примитивных полосок у куриного эмбриона. Curr Top Dev Biol. Актуальные темы биологии развития. 81. С. 135–56. Дои:10.1016 / S0070-2153 (07) 81004-0. ISBN 9780123742537. PMID 18023726.
- ^ Grapin-Botton, A .; Констам, Д. (2007). «Эволюция механизмов и молекулярного контроля образования энтодермы». Механизмы развития. 124 (4): 253–78. Дои:10.1016 / j.mod.2007.01.001. PMID 17307341. S2CID 16552755.
- ^ а б c Тернер, Дэвид А .; Руэ, По; Маккензи, Джонатан П .; Дэвис, Элеонора; Мартинес Ариас, Альфонсо (01.01.2014). «Brachyury взаимодействует с передачей сигналов Wnt / β-катенина, чтобы вызвать поведение, подобное примитивным полосам, при дифференцировке эмбриональных стволовых клеток мыши». BMC Биология. 12: 63. Дои:10.1186 / s12915-014-0063-7. ISSN 1741-7007. ЧВК 4171571. PMID 25115237.
- ^ а б Warmflash, Арье; Сорре, Бенуа; Etoc, Фред; Сиггиа, Эрик Д; Бриванлоу, Али Х (2014). «Метод повторения раннего эмбрионального пространственного паттерна в человеческих эмбриональных стволовых клетках». Методы природы. 11 (8): 847–854. Дои:10.1038 / nmeth.3016. ЧВК 4341966. PMID 24973948.
- ^ а б Etoc, Фред; Мецгер, Якоб; Рузо, Альберт; Кирст, Кристоф; Йони, Анна; Озаир, М. Зишан; Brivanlou, Ali H .; Сиггиа, Эрик Д. (2016). «Баланс между секретируемыми ингибиторами и контролем самоорганизации гаструлоидов». Клетка развития. 39 (3): 302–315. Дои:10.1016 / j.devcel.2016.09.016. ЧВК 5113147. PMID 27746044.
- ^ а б Бринк, Сюзанна К. ван ден; Бэйли-Джонсон, Питер; Балайо, Тина; Хаджантонакис, Анна-Катерина; Новочин, Соня; Тернер, Дэвид А .; Ариас, Альфонсо Мартинес (2014-11-15). «Нарушение симметрии, спецификация зародышевого листка и осевая организация в агрегатах эмбриональных стволовых клеток мыши». Разработка. 141 (22): 4231–4242. Дои:10.1242 / dev.113001. ISSN 0950-1991. ЧВК 4302915. PMID 25371360.
- ^ а б c Тернер, Дэвид Эндрю; Glodowski, Cherise R .; Луз, Алонсо-Крисостомо; Бэйли-Джонсон, Питер; Hayward, Penny C .; Коллиньон, Жером; Густавсен, Карстен; Серуп, Палле; Шретер, Кристиан (13.05.2016). «Взаимодействия между Nodal и Wnt, сигнализирующими об устойчивом нарушении симметрии и осевой организации в гаструлоидах (эмбриональные органоиды)». bioRxiv 10.1101/051722.
- ^ а б c Тернер, Дэвид; Алонсо-Крисостомо, Лус; Гиргин, Мехмет; Бэйли-Джонсон, Питер; Glodowski, Cherise R .; Хейворд, Пенелопа С .; Коллиньон, Жером; Густавсен, Карстен; Серуп, Палле (31.01.2017). «Гаструлоиды развивают три оси тела в отсутствие внеэмбриональных тканей и пространственно локализованной передачи сигналов». bioRxiv 10.1101/104539.
- ^ Беккари, Леонардо; Морис, Наоми; Гиргин, Мехмет; Тернер, Дэвид А .; Бэйли-Джонсон, Питер; Косси, Энн-Катрин; Lutolf, Matthias P .; Дубуль, Дени; Ариас, Альфонсо Мартинес (октябрь 2018 г.). «Многоосные свойства самоорганизации эмбриональных стволовых клеток мыши в гаструлоиды». Природа. 562 (7726): 272–276. Bibcode:2018Натура.562..272Б. Дои:10.1038 / s41586-018-0578-0. ISSN 0028-0836. PMID 30283134. S2CID 52915553.
- ^ Тернер, Дэвид А .; Гиргин, Мехмет; Алонсо-Крисостомо, Лус; Триведи, Викас; Бэйли-Джонсон, Питер; Glodowski, Cherise R .; Хейворд, Пенелопа С .; Коллиньон, Жером; Густавсен, Карстен (2017-11-01). «Переднезадняя полярность и удлинение при отсутствии внеэмбриональных тканей и пространственно локализованной передачи сигналов в гаструлоидах: органоиды эмбрионов млекопитающих». Разработка. 144 (21): 3894–3906. Дои:10.1242 / dev.150391. ISSN 0950-1991. ЧВК 5702072. PMID 28951435.
Библиография
- Арнольд, Себастьян Дж .; Робертсон, Элизабет Дж. (2009). «Взяв на себя обязательство: распределение клеточных клонов и формирование осевого паттерна в раннем эмбрионе мыши». Nat. Преподобный Мол. Cell Biol. 10 (2): 91–103. Дои:10.1038 / nrm2618. PMID 19129791. S2CID 94174.

- Катала, Мартин (2005). «Эмбриология позвоночника и спинного мозга». В Тортори-Донати, Паоло; и другие. (ред.). Детская нейрорадиология: мозг. Springer. ISBN 978-3-540-41077-5.
- Ересковский, Александр В. (2010). Сравнительная эмбриология губок. Springer. ISBN 978-90-481-8574-0.
- Гилберт, Скотт Ф. (2010). Биология развития (Девятое изд.). Sinauer Associates. ISBN 978-0-87893-558-1.
- Холл, Брайан Кейт (1998). «8.3.3 Гаструла и гаструляция». Эволюционная биология развития (2-е изд.). Нидерланды: Kluwer Academic Publishers. ISBN 978-0-412-78580-1.
- Харрисон, Лайонел Г. (2011). Формирование жизни: создание биологической модели. Издательство Кембриджского университета. ISBN 978-0-521-55350-6.
- Макгиди, Томас А., изд. (2006). «Гаструляция». Ветеринарная эмбриология. Вили-Блэквелл. ISBN 978-1-4051-1147-8.
- Мундлос, Стефан (2009). «Действие гена: генетика развития». В Speicher, Майкл; и другие. (ред.). Генетика человека Фогеля и Мотульского: проблемы и подходы (4-е изд.). Springer. Дои:10.1007/978-3-540-37654-5. ISBN 978-3-540-37653-8.
- Там, Патрик П.Л .; Берингер, Ричард Р. (1997). «Мышиная гаструляция: формирование плана тела млекопитающих». Мех. Dev. 68 (1–2): 3–25. Дои:10.1016 / S0925-4773 (97) 00123-8. PMID 9431800. S2CID 14052942.

дальнейшее чтение
- Барон, Маргарет Х. (2001). «Эмбриональная индукция гемопоэза и васкулогенеза млекопитающих». В Зоне, Леонард I. (ред.). Гематопоэз: подход к развитию. Издательство Оксфордского университета. ISBN 978-0-19-512450-7.
- Каллен, К. (2009). «эмбриология и раннее развитие животных». Энциклопедия наук о жизни, Том 2. Информационная база. ISBN 978-0-8160-7008-4.
- Форгач, Г. и Ньюман, Стюарт А. (2005). «Дробление и образование бластулы». Биологическая физика развивающегося эмбриона. Издательство Кембриджского университета. Bibcode:2005bpde.book ..... F. ISBN 978-0-521-78337-8.CS1 maint: использует параметр авторов (связь)
- Форгач, Г. и Ньюман, Стюарт А. (2005). «Эпителиальный морфогенез: гаструляция и нейруляция». Биологическая физика развивающегося эмбриона. Издательство Кембриджского университета. Bibcode:2005bpde.book ..... F. ISBN 978-0-521-78337-8.CS1 maint: использует параметр авторов (связь)
- Харт, Натан Х. и Флак, Ричард А. (1995). «Эпиболия и гаструляция». В Capco, Дэвид (ред.). Цитоскелетные механизмы во время развития животных. Академическая пресса. ISBN 978-0-12-153131-7.CS1 maint: использует параметр авторов (связь)
- Knust, Элизабет (1999). «Гаструляционные движения». В Бирчмайере, Вальтер; Берчмайер, Кармен (ред.). Эпителиальный морфогенез в развитии и заболевании. CRC Press. С. 152–153. ISBN 978-90-5702-419-1.
- Кунц, Иветт В. (2004). «Гаструляция». Биология развития костистых рыб. Springer. ISBN 978-1-4020-2996-7.
- Нация, Джеймс Л., изд. (2009). «Гаструляция». Физиология и биохимия насекомых. CRC Press. ISBN 978-0-8493-1181-9.
- Росс, Лоуренс М .; Ламперти, Эдвард Д., ред. (2006). «Онтогенез человека: гаструляция, невруляция и образование сомитов». Атлас анатомии: общая анатомия и костно-мышечная система. Тиме. ISBN 978-3-13-142081-7.
- Sanes, Dan H. et al. (2006). «Ранняя эмбриология многоклеточных животных». Развитие нервной системы (2-е изд.). Академическая пресса. С. 1–2. ISBN 978-0-12-618621-5.CS1 maint: использует параметр авторов (связь)
- Стангер, Бен З. и Мелтон, Дуглас А. (2004). «Развитие энтодермальных производных в легких, печени, поджелудочной железе и кишечнике». В Эпштейне, Чарльз Дж .; и другие. (ред.). Врожденные ошибки развития: молекулярные основы клинических нарушений морфогенеза. Издательство Оксфордского университета. ISBN 978-0-19-514502-1.CS1 maint: использует параметр авторов (связь)