Модель Морриса – Лекара - Morris–Lecar model
В Модель Морриса – Лекара это модель биологического нейрона разработан Кэтрин Моррис и Гарольдом Лекаром для воспроизведения разнообразия колебательный поведение по отношению к Ca++ и K+ проводимость в мышечном волокне гигантская ракушка .[1] Нейроны Морриса-Лекара проявляют как класс I, так и класс II. нейрон возбудимость.
История
Кэтрин Моррис (род. 24 декабря 1949 г.) - канадский биолог. Она выиграла стипендию Содружества на обучение в Кембриджский университет, где в 1977 году защитила докторскую диссертацию. Она стала профессором Университет Оттавы в начале 1980-х гг. По состоянию на 2015 год она Почетный профессор в Университете Оттавы. Гарольд Лекар (18 октября 1935 г. - 4 февраля 2014 г.) был американским профессором биофизики и нейробиологии в Калифорнийский университет в Беркли. Он окончил со степенью доктора физики Колумбийский университет в 1963 г.
Экспериментальный метод
Эксперименты Морриса-Лекара основывались на методе фиксации тока, установленном Кейнс и другие. (1973).[2]
Крупные экземпляры ракушка Balanus nubilus (Pacific Bio-Marine Laboratories Inc., Венеция, Калифорния). Моллюск распиливали на боковые половины и осторожно обнажали мышцы, отвечающие за депрессор scutorum rostralis. Отдельные волокна рассекались, разрез начинался от сухожилия. Другой конец мышцы разрезали рядом с местом ее прикрепления к оболочке и лигатурировали. Изолированные волокна либо использовались немедленно, либо перед использованием выдерживались до 30 мин в стандартной искусственной морской воде (ASW; см. Ниже). Эксперименты проводились при комнатной температуре 22 C.[1]
Основные предположения, лежащие в основе модели Морриса – Лекара.
Среди основных предположений следующие:
- Уравнения применимы к участку пространственно изопотенциальной мембраны. Существуют два постоянных (не инактивируемых) тока, управляемых напряжением, с противоположно смещенными реверсивными потенциалами. Деполяризующий ток переносится ионами Na + или Ca2 + (или обоими), в зависимости от моделируемой системы, а гиперполяризационный ток переносится K +.
- Активационные вентили достаточно быстро отслеживают изменения мембранного потенциала, так что активирующая проводимость может мгновенно снижаться до своего установившегося значения при любом напряжении.
- Динамика переменной восстановления может быть аппроксимирована линейным дифференциальным уравнением первого порядка для вероятности открытия канала.[3]
Физиологическое описание
Модель Морриса – Лекара представляет собой двумерную систему нелинейный дифференциальные уравнения. Считается упрощенной моделью по сравнению с четырехмерной. Модель Ходжкина – Хаксли.
Качественно эта система уравнений описывает сложную взаимосвязь между мембранным потенциалом и активацией ионных каналов внутри мембраны: потенциал зависит от активности ионных каналов, а активность ионных каналов зависит от напряжения. При изменении параметров бифуркации проявляются разные классы поведения нейронов. τN связана с относительными временными масштабами динамики зажигания, которая широко варьируется от ячейки к ячейке и демонстрирует значительную температурную зависимость.[3]
Количественно:
![{ displaystyle { begin {align} C { frac {dV} {dt}} & ~ = ~ I-g _ { mathrm {L}} (V-V _ { mathrm {L}}) - g _ { mathrm {Ca}} M _ { mathrm {ss}} (V-V _ { mathrm {Ca}}) - g _ { mathrm {K}} N (V-V _ { mathrm {K}}) [ 5pt] { frac {dN} {dt}} & ~ = ~ { frac {N _ { mathrm {ss}} -N} { tau _ {N}}} end {align}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/d26035b4c5fb07ea6ee73a471493a31c1d90e516)
где
![{ displaystyle { begin {align} M _ { mathrm {ss}} & ~ = ~ { frac {1} {2}} cdot left (1+ tanh left [{ frac {V-V_ {1}} {V_ {2}}} right] right) [5pt] N _ { mathrm {ss}} & ~ = ~ { frac {1} {2}} cdot left (1 + tanh left [{ frac {V-V_ {3}} {V_ {4}}} right] right) [5pt] tau _ {N} & ~ = ~ 1 / left ( varphi cosh left [{ frac {V-V_ {3}} {2V_ {4}}} right] right) end {align}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/fa3834e923316f95fbd0e0502324dd4633b4dd3a)
Обратите внимание, что Mсс и Nсс уравнения также могут быть выражены как Mсс = (1 + ехр [−2 (V − V1) / V2])−1 и Nсс = (1 + ехр [−2 (V − V3) / V4])−1, однако большинство авторов предпочитают форму с использованием гиперболических функций.
Переменные
- V : мембранный потенциал
- N : переменная восстановления: вероятность того, что канал K + проводит
Параметры и константы
- я : приложенный ток
- C : емкость мембраны
- гL, гCa, гK : утечка, Ca++, а K+ проводимости через мембранный канал
- VL, VCa, VK : равновесный потенциал соответствующих ионных каналов
- V1, V2, V3, V4: параметры настройки для установившегося режима и постоянной времени
- φ: эталонная частота
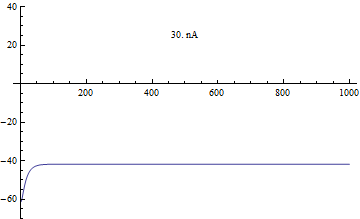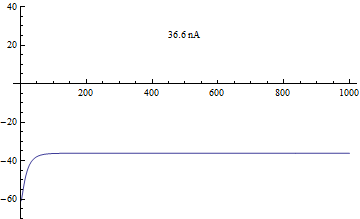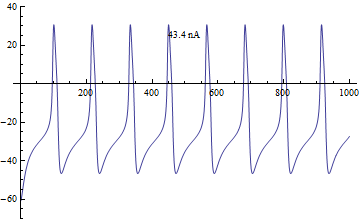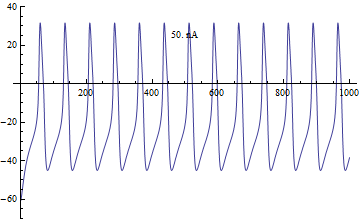
Бифуркации
Бифуркации в модели Морриса – Лекара были проанализированы с приложенным током я, как основной параметр бифуркации и φ, гCa, V3, V4 как второстепенные параметры для фазовая плоскость анализ.[4]


Смотрите также
- Вычислительная нейробиология
- Биологическая модель нейрона
- Модель Ходжкина – Хаксли
- Модель ФитцХью – Нагумо
- Нейронные колебания
- Нелинейная динамика
- Количественные модели потенциала действия
использованная литература
- ^ а б Моррис, Кэтрин; Лекар, Гарольд (июль 1981), «Колебания напряжения в мышечном волокне усоногих гигантских», Биофиз. Дж., 35 (1): 193–213, Bibcode:1981BpJ .... 35..193M, Дои:10.1016 / S0006-3495 (81) 84782-0, ЧВК 1327511, PMID 7260316
- ^ Кейнс, RD; Рохас, Э; Тейлор, RE; Вергара, J (март 1973), «Кальциевая и калиевая системы гигантского мышечного волокна усоногих под контролем мембранного потенциала», Журнал физиологии, 229 (2): 409–455, Дои:10.1113 / jphysiol.1973.sp010146, ЧВК 1350315, PMID 4724831, заархивировано из оригинал на 2013-08-01
- ^ а б Это предположение никогда не бывает точным, поскольку белки каналов состоят из субъединиц, которые должны действовать согласованно, чтобы достичь открытого состояния. Несмотря на отсутствие задержек в начале восстановления, модель кажется адекватной для рассмотрения фазовой плоскости для многих возбудимых систем.Лекар, Гарольд (2007), «Модель Морриса – Лекара», Scholarpedia, 2 (10): 1333, Bibcode:2007SchpJ ... 2.1333л, Дои:10.4249 / scholarpedia.1333
- ^ Цумото, Куничика; Китаджимаб, Хироюки; Йошинагач, Тецуя; Айхарад, Казуюки; Кавакамиф, Хироши (январь 2006 г.), «Бифуркации в модели нейрона Морриса – Лекара» (PDF), Нейрокомпьютинг (на английском и японском языках), 69 (4–6): 293–316, Дои:10.1016 / j.neucom.2005.03.006, заархивировано из оригинал (PDF) на 2012-04-02, получено 2011-09-10
