Количественные модели потенциала действия - Quantitative models of the action potential
В нейрофизиология, несколько математических моделей потенциал действия были разработаны, которые делятся на два основных типа. Первый тип направлен на количественное моделирование экспериментальных данных, то есть точное воспроизведение измерений тока и напряжения. Известный Модель Ходжкина – Хаксли аксона от Лолиго squid служит примером таких моделей.[1] Хотя качественно модель H-H верна, она не описывает точно каждый тип возбудимой мембраны, поскольку она учитывает только два иона (натрий и калий), каждый из которых имеет только один тип чувствительного к напряжению канала. Однако другие ионы, такие как кальций может быть важным, и существует большое разнообразие каналов для всех ионов.[2] Например, потенциал сердечного действия иллюстрирует, как потенциалы действия различной формы могут быть созданы на мембранах с чувствительными к напряжению кальциевыми каналами и различными типами натриевых / калиевых каналов. Второй тип математической модели - это упрощение первого типа; цель состоит не в том, чтобы воспроизвести экспериментальные данные, а в том, чтобы качественно понять роль потенциалов действия в нервных цепях. Для этой цели подробные физиологические модели могут быть излишне сложными и могут скрывать «лес за деревьями». В Модель ФитцХью – Нагумо типичен для этого класса, который часто исследуют на предмет поведение вовлечения.[3] Увлечение обычно наблюдаемый в природе, например при синхронизированном освещении светлячки, который координируется всплеском потенциалов действия;[4] увлечение также можно наблюдать в отдельных нейронах.[5] Оба типа моделей можно использовать для понимания поведения малых биологические нейронные сети, такой как генераторы центральных паттернов отвечает за некоторые автоматические рефлекторные действия.[6] Такие сети могут генерировать сложный временной паттерн потенциалов действия, который используется для координации мышечных сокращений, например, участвующих в дыхании или быстром плавании, чтобы спастись от хищника.[7]
Модель Ходжкина – Хаксли

В 1952 г. Алан Ллойд Ходжкин и Эндрю Хаксли разработали набор уравнений, чтобы соответствовать их экспериментальным данным фиксации напряжения на аксональной мембране.[1][8] Модель предполагает, что емкость мембраны C постоянно; таким образом, трансмембранное напряжение V изменяется с общим трансмембранным током ямалыш согласно уравнению

куда яNa, яK, и яL токи, передаваемые через местные натриевые каналы, калиевые каналы и каналы «утечки» (всеохватывающее), соответственно. Начальный срок ядоб представляет ток, поступающий из внешних источников, таких как возбуждающие постсинаптические потенциалы от дендритов или электрода ученого.
Модель также предполагает, что данный ионный канал либо полностью открыт, либо закрыт; если закрыто, его проводимость равен нулю, тогда как в открытом состоянии его проводимость - некоторая постоянная величина грамм. Следовательно, чистый ток через ионный канал зависит от двух переменных: вероятности поткрыто открытого канала и разницы в напряжении от равновесного напряжения этого иона, V − Vэкв. Например, ток через калиевый канал можно записать как

что эквивалентно Закон Ома. По определению нет чистого тока (яK = 0), когда трансмембранное напряжение равно равновесному напряжению этого иона (когда V = EK).
Чтобы точно соответствовать своим данным, Ходжкин и Хаксли предположили, что каждый тип ионного канала имеет несколько «ворот», так что канал был открыт только в том случае, если все ворота были открыты, а в противном случае - закрыты. Они также предположили, что вероятность открытия ворот не зависит от открытия других ворот; это предположение было позже подтверждено для ворот инактивации.[9] Ходжкин и Хаксли смоделировали чувствительный к напряжению калиевый канал как имеющий четыре затвора; позволяя пп обозначают вероятность того, что один такой вентиль будет открыт, вероятность того, что весь канал будет открыт, является произведением четырех таких вероятностей, т.е. поткрытый, K = п4. Аналогичным образом была смоделирована вероятность того, что чувствительный к напряжению натриевый канал имеет три одинаковых логических элемента вероятности. м и четвертый вентиль, связанный с инактивацией, вероятности час; таким образом, поткрытый, Na = м3час. Предполагается, что вероятности для каждого гейта подчиняются кинетика первого порядка

где как равновесное значение мэкв а постоянная времени релаксации τм зависят от мгновенного напряжения V через мембрану. Если V изменяется во времени медленнее, чем τм, то м вероятность всегда будет примерно равна своему равновесному значению мэкв; однако, если V меняется быстрее, тогда м будет отставать мэкв. Подбирая свои данные фиксации напряжения, Ходжкин и Хаксли смогли смоделировать, как эти равновесные значения и постоянные времени меняются в зависимости от температуры и трансмембранного напряжения.[1] Формулы сложны и экспоненциально зависят от напряжения и температуры. Например, постоянная времени для вероятности активации натриевого канала час изменяется как 3(θ − 6.3) / 10 с температурой Цельсия θ, а с напряжением V в качестве

Таким образом, уравнения Ходжкина – Хаксли являются комплексными, нелинейными. обыкновенные дифференциальные уравнения в четыре независимые переменные: трансмембранное напряжение V, а вероятности м, час и п.[10] Общего решения этих уравнений не найдено. Менее амбициозный, но обычно применимый метод изучения таких нелинейных динамических систем состоит в рассмотрении их поведения в окрестности фиксированная точка.[11] Этот анализ показывает, что система Ходжкина – Хаксли претерпевает переход от стабильного покоя к разрыв колебания как стимулирующий ток ядоб постепенно увеличивается; Примечательно, что аксон снова становится стабильно неподвижным по мере дальнейшего увеличения стимулирующего тока.[12] Также было проведено более общее исследование типов качественного поведения аксонов, предсказываемых уравнениями Ходжкина – Хаксли.[10]
Модель ФитцХью – Нагумо
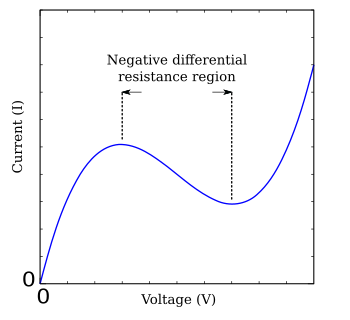
Из-за сложности уравнений Ходжкина – Хаксли были разработаны различные упрощения, которые демонстрируют качественно похожее поведение.[3][13] В Модель ФитцХью – Нагумо является типичным примером такой упрощенной системы.[14][15] На основе туннельный диод, модель FHN имеет только две независимые переменные, но демонстрирует поведение устойчивости, подобное полному уравнению Ходжкина – Хаксли.[16] Уравнения


куда г (В) является функцией напряжения V с областью отрицательного наклона в середине, окруженной одним максимумом и одним минимумом (рисунок FHN). Много изученным простым случаем модели ФитцХью – Нагумо является модель Бонхёффера-ван дер Поля, которая описывается уравнениями[17]


где коэффициент ε предполагается малым. Эти уравнения можно объединить в дифференциальное уравнение второго порядка

Этот ван дер Поль уравнение стимулировало множество исследований в области математики нелинейных динамические системы. Операционный усилитель схемы, реализующие модели потенциала действия FHN и ван дер Поля, были разработаны Кинером.[18]
Гибрид моделей Ходжкина – Хаксли и ФитцХью – Нагумо был разработан Моррисом и Лекаром в 1981 году и применен к мышца волокно ракушки.[19] Верная физиологии ракушек, модель Морриса – Лекара заменяет регулируемый по напряжению натриевый ток модели Ходжкина – Хаксли на зависимый от напряжения кальциевый ток. Нет инактивации (нет час переменная), и кальциевый ток мгновенно уравновешивается, так что снова есть только две переменные, зависящие от времени: трансмембранное напряжение V и вероятность калиевых ворот п. Взрыв, унос и другие математические свойства этой модели были подробно изучены.[20]
Самыми простыми моделями потенциала действия являются модели «промыть и заполнить» (также называемые моделями «интегрировать и запустить»), в которых входной сигнал суммируется (фаза «заполнения») до тех пор, пока не достигнет порога, запускающего импульс и сброс суммирования на ноль (фаза «промывка»).[3][21][22] Все эти модели способны демонстрировать увлечение, который обычно наблюдаемый в нервной системе.[3]
Внеклеточные потенциалы и токи
В то время как приведенные выше модели моделируют трансмембранное напряжение и ток на одном участке мембраны, другие математические модели относятся к напряжениям и токам в ионном растворе, окружающем нейрон.[23] Такие модели полезны при интерпретации данных от внеклеточных электродов, которые были обычным явлением до изобретения стеклянного электрода-пипетки, позволяющего производить внутриклеточную запись.[24] Внеклеточную среду можно смоделировать как нормальную изотропную ионный раствор; в таких решениях ток следует за электрический полевые линии, согласно континуальной форме Закон Ома

куда j и E векторы, представляющие плотность тока и электрическое поле соответственно, и где σ - проводимость. Таким образом, j можно найти из E, который, в свою очередь, можно найти с помощью Уравнения Максвелла. Уравнения Максвелла можно свести к относительно простой задаче электростатика, поскольку концентрации ионов меняются слишком медленно (по сравнению с скорость света ) за магнитные эффекты быть важным. В электрический потенциал φ (Икс) в любой внеклеточной точке Икс можно решить с помощью Личность Грина[23]

где интегрирование происходит по всей поверхности мембраны; позиция на мембране, σвнутри и φвнутри - проводимость и потенциал внутри мембраны, а σза пределами и φза пределами соответствующие значения сразу за пределами мембраны. Таким образом, при этих значениях σ и φ на мембране внеклеточный потенциал φ (Икс) можно рассчитать для любой позиции Икс; в свою очередь, электрическое поле E и плотность тока j можно рассчитать из этого потенциального поля.[25]

Смотрите также
- Биологические модели нейронов
- Текущее уравнение GHK
- Модели нейронных вычислений
- Соляная проводимость
- Биоэлектроника
- Теория кабеля
Рекомендации
- ^ а б c Ходжкин А.Л., Хаксли А.Ф., Кац Б (1952). «Токи, переносимые ионами натрия и калия через мембрану гигантского аксона Лолиго». Журнал физиологии. 116 (4): 424–448. Дои:10.1113 / jphysiol.1952.sp004717. ЧВК 1392213. PMID 14946713.CS1 maint: несколько имен: список авторов (связь)
Ходжкин А.Л., Хаксли А.Ф. (1952). «Токи, переносимые ионами натрия и калия через мембрану гигантского аксона Лолиго». Журнал физиологии. 116 (4): 449–472. Дои:10.1113 / jphysiol.1952.sp004717. ЧВК 1392213. PMID 14946713.
Ходжкин А.Л., Хаксли А.Ф. (1952). «Компоненты проводимости мембраны в гигантском аксоне Лолиго». J Physiol. 116 (4): 473–496. Дои:10.1113 / jphysiol.1952.sp004718. ЧВК 1392209. PMID 14946714.
Ходжкин А.Л., Хаксли А.Ф. (1952). «Двойное влияние мембранного потенциала на проводимость натрия в гигантском аксоне Лолиго». J Physiol. 116 (4): 497–506. Дои:10.1113 / jphysiol.1952.sp004719. ЧВК 1392212. PMID 14946715.
Ходжкин А.Л., Хаксли А.Ф. (1952). «Количественное описание мембранного тока и его применение к проводимости и возбуждению в нерве». J Physiol. 117 (4): 500–544. Дои:10.1113 / jphysiol.1952.sp004764. ЧВК 1392413. PMID 12991237. - ^ Ямада WM, Кох С, Адамс PR (1989). «Множественные каналы и динамика кальция». В К. Кох, Я Сегев (ред.). Методы нейронного моделирования: от синапсов к сетям. Кембридж, Массачусетс: Книга Брэдфорда, MIT Press. С. 97–133. ISBN 978-0-262-11133-1.CS1 maint: несколько имен: список авторов (связь)
- ^ а б c d Хоппенстедт (1986). Введение в математику нейронов. Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-31574-6.
- ^ Hanson, F.E .; Case, J.F .; Buck, E .; Бак, Дж. (1971). "Синхронность и мгновенное вовлечение в светлячок Новой Гвинеи". Наука. 174 (4005): 161–164. Bibcode:1971 г., наука ... 174..161H. Дои:10.1126 / science.174.4005.161. PMID 17742039.
- ^ Гуттман Р., Фельдман Л., Якобссон Э (1980). «Частотный захват аксона кальмара». J. Membr. Биол. 56 (1): 9–18. Дои:10.1007 / BF01869347. PMID 7441721.
- ^ Получение ПА (1989). «Реконструкция малых нейронных сетей». В C Koch и я Сегев (ред.). Методы нейронного моделирования: от синапсов к сетям. Кембридж, Массачусетс: Книга Брэдфорда, MIT Press. С. 171–194. ISBN 978-0-262-11133-1.
- ^ Хупер, Скотт Л. «Генераторы центральных образов». Эмбриональный ELS (1999) http://www.els.net/elsonline/figpage/I0000206.html[постоянная мертвая ссылка ] (2 из 2) [6 февраля 2001 г., 11:42:28] Интернет: доступ 27 ноября 2007 г. [1].
- ^ Нельсон М.Э., Ринзель Дж. (1994). "Модель Ходжкина – Хаксли" (PDF). В Bower J, Beeman D (ред.). Книга GENESIS: Изучение реалистичных нейронных моделей с помощью общей системы нейронной симуляции. Нью-Йорк: Springer Verlag. С. 29–49.
- ^ Армстронг CM, Безанилла Ф, Рохас Э. (1973). «Разрушение инактивации натриевой проводимости в аксонах кальмаров, перфузированных проназой». J. Gen. Physiol. 62 (4): 375–391. Дои:10.1085 / jgp.62.4.375. ЧВК 2226121. PMID 4755846.
Рохас Э., Руди Б. (1976). «Нарушение инактивации натриевой проводимости специфической протеазой в перфузируемых нервных волокнах от Лолиго". J. Physiol. 262 (2): 501–531. Дои:10.1113 / jphysiol.1976.sp011608. ЧВК 1307656. PMID 994046. - ^ а б Сато С, Фукай Х, Номура Т, Дои С (2005). "Бифуркационный анализ уравнений Ходжкина – Хаксли". В Reeke GN, Poznanski RR, Lindsay KA, Rosenberg JR, Sporns O (ред.). Моделирование в нейронауках: от биологических систем до нейромиметической робототехники (2-е изд.). Бока-Ратон: CRC Press. С. 459–478. ISBN 978-0-415-32868-5.
- ^ Guckenheimer J, Холмс П. (1986). Нелинейные колебания, динамические системы и бифуркации векторных полей. (2-е издание, переработанное и исправленное ред.). Нью-Йорк: Springer Verlag. С. 12–16. ISBN 978-0-387-90819-9.
- ^ Сабах Н.Х., Спанглер Р.А. (1970). «Повторяющийся ответ модели Ходжкина-Хаксли для гигантского аксона кальмара». Журнал теоретической биологии. 29 (2): 155–171. Дои:10.1016/0022-5193(70)90017-2. PMID 5500466.
Эванс Дж. В. (1972). «Уравнения нервного аксона. I. Линейные приближения». Indiana Univ. Математика. J. 21 (9): 877–885. Дои:10.1512 / iumj.1972.21.21071.
Эванс Дж. У., Феро Дж. (1977). «Теория локальной устойчивости нервного импульса». Математика. Biosci. 37 (1–2): 23–50. Дои:10.1016/0025-5564(77)90076-1. - ^ ФитцХью Р. (1960). "Пороги и плато в уравнениях нервов Ходжкина-Хаксли". J. Gen. Physiol. 43 (5): 867–896. Дои:10.1085 / jgp.43.5.867. ЧВК 2195039. PMID 13823315.
Кеплер ТБ, Эбботт Л.Ф., Мардер Э. (1992). «Редукция моделей нейронов на основе проводимости». Биологическая кибернетика. 66 (5): 381–387. Дои:10.1007 / BF00197717. PMID 1562643. - ^ ФитцХью Р. (1961). «Импульсы и физиологические состояния в теоретических моделях нервной мембраны». Биофизический журнал. 1 (6): 445–466. Bibcode:1961BpJ ..... 1..445F. Дои:10.1016 / S0006-3495 (61) 86902-6. ЧВК 1366333. PMID 19431309.
- ^ Нагумо Дж., Аримото С., Йошизава С. (1962). «Активная линия передачи импульсов, имитирующая аксон нерва». Труды IRE. 50 (10): 2061–2070. Дои:10.1109 / JRPROC.1962.288235.
- ^ ФитцХью Р. (1969). «Математические модели возбуждения и распространения в нерве». В HP Schwann (ред.). Биологическая инженерия. Нью-Йорк: Макгроу-Хилл. С. 1–85.
- ^ Бонхёффер К.Ф. (1948). «АКТИВАЦИЯ ПАССИВНОГО ЖЕЛЕЗА КАК МОДЕЛЬ ВОЗБУЖДЕНИЯ НЕРВА». J. Gen. Physiol. 32 (1): 69–91. Дои:10.1085 / jgp.32.1.69. ЧВК 2213747. PMID 18885679.
Бонхёффер К.Ф. (1953). "Modelle der Nervenerregung". Naturwissenschaften. 40 (11): 301–311. Bibcode:1953NW ..... 40..301B. Дои:10.1007 / BF00632438.
ван дер Поль Б (1926). «О релаксационных колебаниях». Философский журнал. 2: 978–992.
ван дер Поль Б, ван дер Марк Дж (1928). «Сердцебиение рассматривается как расслабляющее колебание и электрическая модель сердца». Философский журнал. 6: 763–775.
ван дер Поль Б, ван дер Марк Дж (1929). «Сердцебиение рассматривается как расслабляющее колебание и электрическая модель сердца». Arch. Neerl. Физиол. 14: 418–443. - ^ Кинер Дж. П. (1983). «Аналоговая схема для уравнений Ван дер Поля и ФитцХью-Нагумо». IEEE Transactions по системам, человеку и кибернетике. 13 (5): 1010–1014. Дои:10.1109 / TSMC.1983.6313098.
- ^ Моррис С., Лекар Х (1981). «Колебания напряжения в мышечном волокне усоногих гигантских». Биофизический журнал. 35 (1): 193–213. Bibcode:1981BpJ .... 35..193M. Дои:10.1016 / S0006-3495 (81) 84782-0. ЧВК 1327511. PMID 7260316.
- ^ Ринзель Дж., Эрментрут Г.Б. (1989). «Анализ нервной возбудимости и колебаний». В К. Кох, Я Сегев (ред.). Методы нейронного моделирования: от синапсов к сетям. Кембридж, Массачусетс: Книга Брэдфорда, MIT Press. С. 135–169. ISBN 978-0-262-11133-1.
- ^ Кинер Дж. П., Хоппенстедт ФК, Ринзель Дж. (1981). «Интегрируй и запускай модели реакции нервной мембраны на колебательный вход». Журнал SIAM по прикладной математике. 41 (3): 503–517. Дои:10.1137/0141042.
- ^ Цейнар, Павел; Вышата, Олдржих; Кукал, Яромир; Беранек, Мартин; Валиш, Мартин; Прохазка, Алеш (2020). «Простая модель возбуждающего и тормозящего нейрона с переключателем между конденсаторами и биологически объясненными всеми частями позволяет вводить хаотические колебания, зависящие от схемы зажигания». Научные отчеты. 10 (1): 7353. Дои:10.1038 / s41598-020-63834-7. PMID 32355185.
- ^ а б Стивенс, Чарльз Ф. (1966). Нейрофизиология: учебник. Нью-Йорк: Джон Уайли и сыновья. стр.161 –173. LCCN 66015872.
- ^ Линг G, Джерард RW (1949). «Нормальный мембранный потенциал волокон портняжника лягушки». J. Cell. Комп. Физиол. 34 (3): 383–396. Дои:10.1002 / jcp.1030340304. PMID 15410483.
- ^ Лоренте де Но Р. (1947). «Исследование физиологии нервов». Stud. Rockefeller Inst. Med. Исследование. 132: Гл. 16.
Мауро А. (1960). «Свойства тонких генераторов электрофизиологических потенциалов в объемных проводниках». J. Neurophysiol. 23 (2): 132–143. Дои:10.1152 / jn.1960.23.2.132.
Вудбери JW (1965). «Глава 3: Потенциалы в объемном проводнике». В ТЦ Ручь; HD Паттон (ред.). Физиология и биофизика. Филадельфия: W. B. Saunders Co.
дальнейшее чтение
- Гласс Л., Макки М.С. (1988). От часов к хаосу: ритмы жизни. Принстон, Нью-Джерси: Принстонский университет. ISBN 978-0-691-08496-1.