Hartebeest - Hartebeest
| Hartebeest | |
|---|---|
 | |
| Хартбест кока-колы в Национальный парк Серенгети, Танзания | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Парнокопытные |
| Семья: | Bovidae |
| Подсемейство: | Alcelaphinae |
| Род: | Alcelaphus |
| Разновидность: | A. buselaphus |
| Биномиальное имя | |
| Alcelaphus buselaphus Паллада, 1766 | |
| Подвиды[2] | |
Список
| |
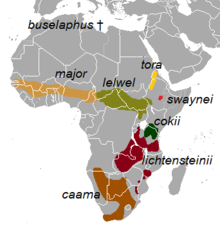 | |
| Распространение подвида | |
| Синонимы[2] | |
| |

В Hartebeest (/ˈчасɑːrтəˌбяsт/;[3] Alcelaphus buselaphus), также известный как Конгони, является Африканский антилопа. 8 подвид были описаны, в том числе два, которые иногда считались независимыми. Большая антилопа, хищник высотой чуть более 1 м (3,3 фута) в плече и имеет типичную длину головы и тела от 200 до 250 см (от 79 до 98 дюймов). Вес составляет от 100 до 200 кг (от 220 до 440 фунтов). У него особенно удлиненный лоб, рога необычной формы, короткая шея и заостренные уши. Его ноги, которые часто имеют черные отметины, необычно длинные. Шерсть обычно короткая и блестящая. Цвет шерсти варьируется в зависимости от подвида, от песочно-коричневого у западного хартбиста до шоколадно-коричневого у хартбиста Суэйна. У обоих полов всех подвидов есть рога, причем у самок они более тонкие. Рога могут достигать длины 45–70 см (18–28 дюймов). Помимо вытянутого лица, большая грудь и резко наклонная спина отличают хартбиста от других антилоп.
Стачные животные, хищники образуют стада от 20 до 300 особей. Они очень бдительны и неагрессивны. В первую очередь они пасутся на траву, и их рацион состоит в основном из трав. Спаривание у hartebeest происходит в течение года с одним или двумя пиками и зависит от подвида и местных факторов. И мужчины, и женщины достигают половая зрелость в возрасте от одного до двух лет. Беременность длится восемь-девять месяцев, после чего рождается один теленок. Пик рождаемости обычно приходится на сухой сезон. Продолжительность жизни от 12 до 15 лет.
Обитает в сухих саванны и лесистые пастбища, хищники часто перемещаются в более засушливые места после дождя. О них сообщалось с высоты на Гора Кения до 4000 м (13000 футов). Хартебист раньше был широко распространен в Африке, но популяция резко сократилась из-за разрушения среды обитания, охоты, расселения людей и конкуренции с домашним скотом за еду. Каждый из восьми подвидов хищника имеет различный природоохранный статус. В Bubal hartebeest был объявлен вымерший посредством Международный союз охраны природы (МСОП) в 1994 году. В то время как популяция красных хищников растет, популяция хищников Тора уже Находящихся под угрозой исчезновения, падают. Hartebeest вымерли в Алжире, Египте, Лесото, Ливии, Марокко, Сомали и Тунисе; но был представил в Свазиленд и Зимбабве. Это популярный игровое животное из-за высоко ценимого мяса.
Этимология
Народное название «hartebeest», возможно, произошло от устаревшего нидерландский язык слово Hertebeest,[4] в прямом смысле олень зверь,[3] основанный на сходстве (с ранними голландскими поселенцами) антилопы с олень.[5] Слово «hartebeest» впервые было использовано в южноафриканской литературе в голландском колониальном администраторе. Ян ван Рибек журнал Дагрегистр в 1660 году. Он писал: «Мистер Питер ein hart-beest geschooten hadde (Мастер Питер [ван Меерхофф] застрелил одного хищника) ».[6] Другое название зверобоя - Конгони,[7] а суахили слово.[8] Kongoni часто используется для обозначения, в частности, одного из подвид —Хартбест кока-колы.[9]
Таксономия
В научное название Hartebeest - это Alcelaphus buselaphus. Первый описанный немецким зоологом Питер Саймон Паллас в 1766 г. отнесен к роду Alcelaphus и помещен в семью Bovidae.[2] В 1979 г. палеонтолог Элизабет Врба поддержанный Sigmoceros как отдельный род для Хищник Лихтенштейна, своего рода хищник, как она предполагала, связано с Connochaetes (антилопа гну).[10][11] Она проанализировала черты черепов живых и вымерших видов антилоп, чтобы сделать кладограмму, и утверждала, что широкий череп связывает зверя Лихтенштейна с Connochaetes.[12] Однако это открытие не было воспроизведено Аланом В. Джентри из Музей естественной истории, которые классифицировали его как самостоятельный вид Alcelaphus.[13] Зоологи, такие как Джонатан Кингдон и Теодор Халтенорт считается подвидом A. buselaphus.[2] Врба распустил новый род в 1997 году после пересмотра.[14] An МтДНК анализ не смог найти никаких доказательств в поддержку отдельного рода хищников Лихтенштейна. Это также показало племя Alcelaphini быть монофилетический, и обнаружил близкое родство между Alcelaphus и дерзкие (род Дамалиск ) - как генетически, так и морфологически.[15]
Подвиды

Выявлено восемь подвидов, из которых два - А. б. каама и А. б. Lichtensteinii - считались самостоятельными видами. Однако генетическое исследование П. Арктандера, проведенное в 1999 г. Копенгагенский университет и коллеги, которые пробовали область контроля из митохондриальная ДНК, обнаружил, что эти двое образовали кладу внутри A. buselaphus, и что признание их за виды сделало бы A. buselaphus парафилетический (неестественная группировка). То же исследование показало А. б. основной быть наиболее расходящимися, разветвившись до разделения родословной, чтобы дать объединенный caama / lichtensteinii родословная и еще одна, которая дала начало оставшимся сохранившийся подвид.[16] Напротив 2001 филогенетический исследование, основанное на D-петля и цитохром b анализ Øystein Flagstad (Норвежский институт исследований природы, Тронхейм ) и его коллеги обнаружили, что южная линия А. б. каама и A. lichtensteinii расходились раньше.[11] Анализ строения черепа поддерживает разделение на три основных раздела: А. б. Buselaphus разделение (назначить, включая также А. б. основной разделение), А. б. тора деление (также включая А. б. Cokii и А. б. Swaynei) и А. б. лелвел разделение.[2] Другой анализ данных последовательностей цитохрома b и D-петли показывает заметное близость между А. б. лелвел и А. б. тора подразделения.[17]
Восемь подвидов, включая два спорных:[1][18]
- † А. б. Buselaphus (Паллас, 1766 г.) : Известный как Bubal hartebeest или северный хартбист. Ранее происходил в Северной Африке, от Марокко к Египет. Он был истреблен к 1920-м годам.[19] Он был объявлен вымершим в 1994 г. Международный союз охраны природы и природных ресурсов (МСОП).[20][21]
- А. б. каама (Saint-Hilaire, 1803) : Известный как красный hartebeest или мыс хартбест. Ранее происходило на юге Ангола; северные и восточные саванны Намибия; центральный, южный и юго-западный Ботсвана; Северный мыс, Восточный мыс, Western Cape, Свободный штат, Северо-Запад и Гаутенг провинции и западные Квазулу-Натал Южной Африки. В настоящее время ликвидированы все эти районы, кроме Северного мыса, центральной и юго-западной Ботсваны и Намибии. В этих странах произошли крупные повторные интродукции.[19] Популяция этого зверобоя постоянно растет.[22]
- А. б. Cokii (Гюнтер, 1884): Известный как Хартбест кока-колы или конгони. Родной и заключенный внутри Кения и северный Танзания.[19]
- А. б. лелвел (Heuglin, 1877) : Известный как Lelwel hartebeest. Ранее находили на севере и северо-востоке. Демократическая Республика Конго; юго-восток и юго-запад Судан; и северо-западная крайность Танзания.[19] Резкое сокращение популяции с 1980-х гг. Привело к тому, что большинство особей было ограничено охраняемыми территориями внутри и за пределами ареала.[23]
- А. б. Lichtensteinii (Питерс, 1849) : Известный как Хищник Лихтенштейна. Обитает в Miombo Woodlands восточной и южной Африки.[24] Родом из Анголы, Демократической Республики Конго, Малави, Мозамбик, Южная Африка, Танзания, Замбия и Зимбабве.[25]
- А. б. основной (Блит, 1869) : Известный как западный hartebeest. Ранее широко распространено в Мали, Нигер, Сенегал, Гамбия, Гвинея-Бисау, Гвинея, Кот-д'Ивуар, Гана, Нигерия, юго-запад Чада, Камерун, запад Центральноафриканской Республики и Бенин. В настоящее время он встречается в гораздо меньших количествах, в основном на охраняемых территориях этих стран. Вероятно, в Гамбии он вымер.[19]
- А. б. Swaynei (Склейтер, 1892) : Известный как Житель Суэйна. Ограничено южным Рифтовая долина в Эфиопии. Раньше это происходило по всей Рифтовой долине, а его ареал простирался с востока на северо-запад. Сомали. К 1930 году он исчез из Сомали.[19] Его популяция очень мала и сокращается.[26]
- А. б. тора (серый, 1873) : Известный как Тора hartebeest. Ранее встречались на северо-западе Эфиопии, а также на западе и юго-западе. Эритрея.[27] Его нынешний статус неясен, хотя местные жители сообщают о небольшом количестве из этих районов.[19]
- Пять подвидов хищников

Bubal hartebeest

Красный hartebeest

Хартбест кока-колы

Хищник Лихтенштейна

Житель Суэйна




Генетика и гибриды

В 2000 году в ходе исследования были изучены две основные популяции хищников Суэйнов: Заповедник дикой природы Сенкеле и Национальный парк Нечисар, для митохондриальной (D-петля) и ядерной (микроспутник ) изменчивость в попытке оценить уровни генетическая вариация между популяциями и внутри подвида. Результаты показали заметную дифференциацию между двумя популяциями; что из заповедника дикой природы Сенкеле показал больше генетическое разнообразие чем из национального парка Нечисар. Еще одним откровением было то, что перемещение особей из заповедника дикой природы Сенкеле в 1974 году не внесли значительного вклада в генофонд национального парка Нечисар. Кроме того, популяции свейнов-хищников сравнивали с большой популяцией красных хищных, и было обнаружено, что оба подвида имеют высокую степень генетической изменчивости. В исследовании высказывались сохранение на месте зверя свейна и возобновленная попытка его перемещения с целью сохранения генетического разнообразия и увеличения его популяции в обеих охраняемых зонах.[17]
В диплоид количество хромосом у хищника составляет 40. Гибриды обычно регистрируются из районов, где ареалы двух подвидов перекрываются.[7] Гибриды между Lelwel и Tora hartebeest были зарегистрированы в восточном Судане и западной Эфиопии, на участке к югу от Голубой Нил примерно до 9 ° северной широты.[28] Исследование подтвердило мужской гибрид красного хищника и блесбок (Damaliscus pygargus) быть стерильный. Стерильность гибрида объяснялась трудностями в сегрегация в течение мейоз, указано азооспермия и небольшое количество стволовые клетки в его семенные канальцы.[29]

Между подвидами существует три четко определенных гибрида:
- Alcelaphus lelwel x cokii: Известен как хартбест Кенийского нагорья или хартбест Лайкипия. Это помесь Lelwel и кока-колы.[30] Этот гибрид светлее по цвету и крупнее, чем хищник кока-колы. Это светлый бугристый цвет с красновато-коричневой верхней частью, а голова длиннее, чем у Hartebeest кока-колы. У обоих полов есть рога, которые тяжелее и длиннее, чем у родителей. Раньше он был распространен на западном Кенийском нагорье, между Озеро Виктория и Гора Кения, но теперь считается, что она ограничена долиной Ламбве (юго-запад Кении) и Лайкипия и близлежащие регионы западно-центральной Кении.[31][32]
- Жароплодник Джексона не имеет четкого таксономического статуса. Как и форма выше, он считается гибридом между Lelwel и Coke's hartebeest и имеет аналогичное распространение. В База данных африканских антилоп (1998) рассматривает его как синоним хартбеста Lelwel.[19] От озера Баринго до горы Кения хищник Джексона значительно напоминает хищника Лелвела, в то время как от озера Виктория до южной части Рифт-Валли он больше похож на хита кока-колы.[33]
- Alcelaphus lelwel x swaynei : Также известен как хищник Ноймана, названный в честь путешественника и охотника. Артур Генри Нойман.[33] Считается, что это помесь зверя Лелвела и зверя Суэйна.[30] Лицо длиннее, чем у зверя Суэйна. Цвет шерсти - золотисто-коричневый, более светлый к низу. Подбородок имеет оттенок черного, а хвост оканчивается черным пучком. У обоих полов рога длиннее, чем у свиней Суэйнов. Рога растут в форме широкой буквы «V», в отличие от формы широкой скобки у хартебиста Суэйна и узкой буквы «V» у хартбиста Лелвела, изгибаясь назад и немного внутрь. Встречается в Эфиопии, на небольшой территории к востоку от г. Река Омо и к северу от Озеро Туркана, простираясь к северо-востоку от Озеро Чу Бахир чтобы рядом Озеро Чамо.[34]
Эволюция
Род Alcelaphus возник около 4,4 миллиона лет назад в клады чьи другие члены были Damalops, Нумидокапра, Rabaticeras, Мегалотрагус, Ореонагор, и Connochaetes. Анализ с использованием филогеографический закономерности в популяциях хищников предполагали возможное происхождение Alcelaphus в восточной Африке.[35] Alcelaphus быстро излученный через африканские саванны, заменив несколько предыдущих форм (например, родственник Hirola ). Флагстад и его коллеги показали раннее разделение популяций хищных насекомых на две отдельные линии примерно 0,5 миллиона лет назад - одна к северу, а другая к югу от экватора. Северная линия потомков далее разделилась на восточную и западную, почти 0,4 миллиона лет назад, скорее всего, в результате расширения тропические леса Центральной Африки пояс и последующее сжатие саванна местообитания в период глобального потепления. Восточная линия дала начало Hartebeest Кока-колы, Суэйна, Торы и Лелвела; и от западной линии произошли бубал и западный хартбист. Южное происхождение дало начало Лихтенштейну и красному хищнику. Эти два таксона филогенетически близки, разошлись всего 0,2 миллиона лет назад. Исследование пришло к выводу, что эти важные события на протяжении всей эволюции хищников сильно связаны с климатическими факторами и что произошли последовательные вспышки радиации от более постоянной популяции - a рефугиум - в Восточной Африке; это может иметь жизненно важное значение для понимания эволюционной истории не только хищников, но и других млекопитающих африканской саванны.[11]
Самая ранняя летопись окаменелостей датируется около 0,7 миллиона лет назад.[7] Окаменелости красный hartebeest были найдены в Эландсфонтейне, Корнелия (Фри Стейт) и Флорисбад в Южная Африка, а также в Кабве в Замбия.[36] В Израиль, останки зверобоя найдены в северной Негев, Шефела, Sharon Plain и Тель Лахиш. Эта популяция зверобоя первоначально была ограничена открытой местностью самых южных регионов южной Левант. Вероятно, на него охотились в Египте, что повлияло на численность в Леванте и отключило его от основной популяции в Африке.[37]
Описание

Большая антилопа с особенно удлиненным лбом и рогами необычной формы, хищник стоит чуть более 1 м (3,3 фута) в плече и имеет типичную длину головы и тела от 200 до 250 см (от 79 до 98 дюймов). Вес составляет от 100 до 200 кг (от 220 до 440 фунтов). Хвост длиной от 40 до 60 см (от 16 до 24 дюймов) заканчивается черным пучком.[38] Другими отличительными чертами этого хищника являются длинные ноги (часто с черными отметинами), короткая шея и заостренные уши.[39] Исследование коррелировало размер видов хищных насекомых с продуктивность среды обитания и осадки.[40] Западный хартбист - самый крупный подвид с характерной белой линией между глазами.[41] Красный хартбист также крупный, с черным лбом и контрастной светлой полосой между глазами.[42] Большой хищник Lelwel имеет темные полосы на передней части ног.[28] Hartebeest кока-колы умеренно крупный, с более коротким лбом и более длинным хвостом по сравнению с другими подвидами.[43] Лихтенштейн меньшего размера, с темными полосами на передней части ног, как и у Lelwel hartebeest.[44] Hartebeest Swayne меньше, чем Hartebeest Tora, но оба имеют более короткий лоб и похожий внешний вид.[45]
Обычно короткая и блестящая шерсть различается по цвету в зависимости от подвида.[46] Западный хартбист бледно-песочно-коричневый, но передние лапы более темные.[41] Красный hartebeest - красновато-коричневый с темным лицом. Черные отметины можно увидеть на подбородке, задней части шеи, плечах, бедрах и ногах; они резко контрастируют с широкими белыми пятнами, отмечающими его бока и нижнюю часть крупа.[42][47] Lelwel hartebeest имеет красноватый загар.[28] Жареный кока-кола от красноватого до желтовато-коричневого в верхней части, но имеет относительно более легкие ножки и крупу.[43] Герб Лихтенштейна красновато-коричневый, хотя бока имеют более светлый загар, а крупец беловатый.[44] Tora hartebeest имеет темно-красновато-коричневый цвет в верхней части тела, лице, передних лапах и крупе, но задние лапы и низ живота желтовато-белые.[27][48] Hartebeest Swayne - насыщенный шоколадно-коричневый цвет с мелкими белыми пятнами, которые на самом деле являются белыми кончиками его волосков. Лицо у него черное, за исключением шоколадной полоски под глазами. Плечи и верхняя часть ног черные.[45] Волосы на теле хищника имеют мелкую текстуру и имеют длину около 25 мм (1 дюйм).[10] У hartebeest есть предглазничные железы (железы возле глаз) с центральным протоком, которые выделяют темную липкую жидкость у хищников кока-колы и Лихтенштейна, а также бесцветную жидкость у хищников Лелвеля.[46]

У обоих полов всех подвидов есть рога, причем у самок они более тонкие. Рога могут достигать длины 45–70 см (18–28 дюймов); максимальная длина рога составляет 74,9 см (29,5 дюйма), что зарегистрировано у намибийского красного хищника.[38] Рога западного хартбиста толстые и кажутся U-образными спереди и Z-образными с боков, сначала растут назад, а затем вперед, заканчивая резким поворотом назад.[41] Рога красного и лелвельского хищника похожи на рога западного хищника, но при взгляде спереди выглядят V-образными.[28][42] У лихтенштейна толстые параллельные кольчатые рога с плоским основанием. Его рога короче, чем у других подвидов, изогнутые вверх, затем резко вперед, за которыми следует поворот внутрь под углом примерно 45 ° и последний поворот назад.[44] Рога хищника Суэйна тонкие и имеют форму круглых скобок, загибающихся вверх, а затем назад.[45] Рога хищника Тора особенно тонкие и расставлены в стороны, расходясь больше, чем у любого другого подвида.[48]
Помимо вытянутого лица, большая грудь и резко наклонная спина отличают хартбиста от других антилоп.[4] Hartebeest имеет несколько общих физических черт с сассаби (род. Дамалиск), такие как удлиненное и узкое лицо, форма рогов, шерсть текстура и цвет, а также концевой пучок хвоста. В антилопа гну имеют более специализированные черты черепа и рогов, чем хищники.[46] Выставки hartebeest половой диморфизм, но незначительно, так как оба пола несут рога и имеют схожую массу тела. Степень полового диморфизма зависит от подвида. Самцы на 8% тяжелее самок у хищников Суэйна и Лихтенштейна и на 23% тяжелее у красных хищников. В одном исследовании самый высокий диморфизм был обнаружен в массе черепа.[49] Другое исследование пришло к выводу, что продолжительность периода размножения является хорошим предиктором диморфизма ножек (костных структур, из которых растут рога), высоты и веса черепа, а также лучшим показателем окружности рогов.[50]
Экология и поведение
Активны в основном в дневное время, они пасутся рано утром и ближе к вечеру, а около полудня отдыхают в тени. Общительный, вид формирует стада до 300 особей. В местах с обильной травой собирается большее количество. В 1963 году на равнинах недалеко от Секома Пан в Ботсване было зарегистрировано скопление из 10 000 животных.[46] Однако движущиеся стада не так сплочены и часто расходятся. Члены стада можно разделить на четыре группы: территориальные взрослые самцы, нетерриториальные взрослые самцы, молодые самцы и самки с молодыми. Самки образуют группы от пяти до двенадцати особей, в которых насчитывается четыре поколения молодняка. Самки борются за господство над стадом.[38] Спарринг между мужчинами и женщинами является обычным явлением.[7] В возрасте трех-четырех лет самцы могут попытаться взять на себя территория и его женщины-члены. Постоянный самец защищает свою территорию и будет драться, если его спровоцировать.[49] Самец отмечает границу своей территории посредством дефекации.[38]

Hartebeest - удивительно бдительные и осторожные животные с очень развитым животным. мозги.[51][52] Обычно спокойный по природе, хищник может быть свирепым, когда его провоцируют. Во время кормления один человек остается в поисках опасности, часто стоя на термит насыпь, чтобы увидеть дальше. В случае опасности все стадо сбегает одной цепью после того, как особь внезапно уходит.[38] Львы охотятся на взрослых хищников, леопарды, гиены и Дикая собака; гепарды и шакалы цель молодые.[38] Крокодилы также могут охотиться на хищников.[53]
Тонкие длинные лапы хищника позволяют быстро убежать в открытую среду обитания; в случае нападения грозные рога используются для отпугивания хищника. Поднятое положение глаз позволяет животному постоянно осматривать свое окружение, даже когда оно пасется. Морда эволюционировала таким образом, чтобы получать максимум питания даже от скромной диеты.[7] Рога также используются во время боев среди самцов за господство в сезон размножения;[50] грохот рогов достаточно громкий, чтобы его можно было услышать за сотни метров.[7] Начало боя отмечается серией движений головой и стоек, а также нанесением помета на кучу навоза. Противники падают на колени и после удара молотком начинают борьбу, сцепляя рога. Один пытается отбросить голову другого в сторону, чтобы нанести удар рогами в шею и плечи.[49] Ссоры редко бывают серьезными, но могут быть фатальными.[46]
Как и sassabies, hartebeest издают тихие крякающие и хрюкающие звуки. Подростки, как правило, громче взрослых и крякают, когда их тревожат или преследуют.[38] Hartebeest использует дефекацию как обонятельный и визуальный дисплей.[46] Стада обычно ведут оседлый образ жизни и мигрируют только в неблагоприятных условиях, таких как стихийные бедствия.[54] Hartebeest - наименее мигрирующий в племя Alcelaphini (который также включает антилоп гну и сассаби), а также потребляет наименьшее количество воды и имеет наименьшее количество воды. скорость метаболизма среди членов племени.[46]
Паразиты и болезни
Несколько паразиты были изолированы от хищника.[55][56] Эти паразиты регулярно чередуются между хищниками и газелями или гну.[57] Hartebeest может быть заражен тейлериоз из-за Rhipicephalus evertsi и Theileria разновидность.[58] К югу от Сахара, общие паразиты включают Loewioestrus Varolosus, Gedoelstia cristata и Г. хасслери. Последние два вида могут вызывать серьезные заболевания, такие как энцефалит.[59] Однако не всегда паразиты вредны - 252 личинки были найдены в голове одного замбийского человека без каких-либо патогенность.[56] Нематоды, цестоды, парамфистомы; и аскарида Setaria labiatopapillosa также были изолированы от хищника.[60][61] В 1931 году красный хищник в г. Gobabis (Юго-Западная Африка) была заражена длинными тонкими червями. Они были названы Longistrongylus meyeri по их коллекционеру Т. Мейеру.[62]

Рацион питания
Hartebeest - это в первую очередь травоядные, и их рацион в основном состоит из трав.[63] Исследование в Ранчо Nazinga Game в Буркина-Фасо обнаружили, что структура черепа хищника облегчает переваривание и пережевывание продуктов с высоким содержанием клетчатки.[64] Hartebeest потребляет гораздо меньше пищи, чем другие представители Alcelaphini. Длинный тонкий морда Hartebeest помогает питаться листовыми пластинками невысоких трав и отрывать влагалище от стеблей травы. Кроме того, он может получать питательную пищу даже из высоких старческих трав. Эти приспособления жука позволяют животному хорошо кормиться даже в сухой сезон, который обычно является трудным периодом для травоядных.[7] Например, по сравнению с чалая антилопа, зверобоя лучше добывают и пережевывают скудные отростки многолетние травы временами, когда кормов меньше всего.[64] Эти уникальные способности могли позволить хищнику преобладать над другими животными миллионы лет назад, что привело к его успешному облучению в Африке.[7]
Травы обычно составляют не менее 80 процентов рациона хищников, но они составляют более 95 процентов их пищи в сезон дождей, с октября по май. Jasminum kerstingii является частью рациона хищника в начале сезона дождей. В межсезонье они в основном питаются стебли трав.[64] Исследование показало, что хищник способен переваривать большую часть пищи, чем топи и антилопа гну.[65] В районах с дефицитом воды он может питаться дынями, корнями и клубнями.[46]
При исследовании селективности травы среди антилоп гну, зебры и оленя кока-колы, олень показал самую высокую избирательность. Все животные предпочитают Themeda triandra над Pennisetum mezianum и Digitaria macroblephara. В сухой сезон съедалось больше видов трав, чем во влажный.[66]
Размножение

Спаривание у hartebeest происходит в течение года, с одним или двумя пиками, на которые может повлиять доступность пищи.[63] И мужчины, и женщины достигают половая зрелость в возрасте от одного до двух лет. Размножение зависит от подвида и местных факторов.[10] Спаривание происходит на территориях, защищаемых одним самцом, в основном на открытых участках.[63] Самцы могут отчаянно бороться за господство,[49] после чего доминирующий самец нюхает гениталии самки и следует за ней, если она в эструс. Иногда самка во время эструса слегка вытягивает хвост, чтобы сигнализировать о своей восприимчивости.[46] и самец пытается преградить путь женщине. В конце концов, она может остановиться и позволить мужчине сесть на нее. Копуляция непродолжительна и часто повторяется, иногда дважды или чаще в минуту.[46] Любого злоумышленника в это время прогоняют.[38] В больших стадах самки часто спариваются с несколькими самцами.[46]
Беременность длится от восьми до девяти месяцев, после чего рождается один теленок весом около 9 кг (20 фунтов). Пик рождаемости обычно приходится на сухой сезон и происходит в зарослях - в отличие от антилоп гну, которые рожают группами на равнинах.[46] Хотя телята могут передвигаться самостоятельно вскоре после рождения, обычно они лежат на открытом воздухе в непосредственной близости от своих матерей.[30] Теленок отлучен от груди в четыре месяца,[30] но молодые самцы остаются со своими матерями два с половиной года, дольше, чем у других Alcelaphini.[46] Часто уровень смертности молодых самцов высок, так как им приходится сталкиваться с агрессией территориальных взрослых самцов, а также лишать их хорошего корма.[38] Продолжительность жизни от 12 до 15 лет.[63]
Среда обитания
Hartebeest населяют сухие саванны, открытые равнины и лесные луга,[10] часто переезжают в более засушливые места после дождя. Они более терпимы к лесным массивам, чем другие Alcelaphini, и часто встречаются на окраинах лесов.[63] О них сообщалось с высоты на Гора Кения до 4000 м (13000 футов).[1] Известно, что красный хищник перемещается по обширным территориям, а самки бродят по домашним участкам более 1000 км.2 (390 квадратных миль), с мужскими территориями 200 км2 (77 кв. Миль).[67] Женщины в Национальный парк Найроби (Кения) имеют индивидуальные домашние полигоны протяженностью 3,7–5,5 км.2 (1,4–2,1 кв. Мили), которые не связаны с какой-либо одной женской группой. Средние женские ареалы достаточно велики, чтобы включать от 20 до 30 мужских территорий.[39]
Статус и сохранение



Каждый подвид Hartebeest занесен в список МСОП под различным природоохранным статусом. Вид в целом классифицируется как Наименьшие опасения МСОП.[1] Hartebeest вымерли в Алжире, Египте, Лесото, Ливии, Марокко, Сомали и Тунисе.[1]
- В Bubal hartebeest был объявлен вымерший с 1994 г.[20] Немецкий исследователь Генрих Барт в своих работах 1857 года среди причин сокращения их численности называет огнестрельное оружие и европейское вторжение.[68] Он вымер в Тунис к концу 19 века.[69] Последний человек был застрелен Миссур (Алжир ) в 1925 году.[70]
- Хартбест кока-колы внесен в список «Наименее вызывающий беспокойство». Этот вид сильно пострадал от разрушения среды обитания, и сегодня в Мара, Национальный парк Серенгети, и Национальный парк Тарангире в Танзании и Национальный парк Цаво Восток в Кении. Население сокращается, и 70% населения проживает на охраняемых территориях.[71]
- В Lelwel hartebeest занесен в список находящихся под угрозой исчезновения, и их численность значительно снизилась с 1980-х годов, когда его население составляло более 285 000 человек. Ранее он был распространен в основном в Центральноафриканской Республике, Эфиопии, на севере и северо-востоке Демократической Республики Конго и на юге Судана.[19] Осталось менее 70 000 человек.[23] Большая часть населения в настоящее время проживает в Чаде в Саламатский район и Национальный парк Закума (Чад), население национального парка, пользующееся улучшенной защитой и наблюдающее рост населения с 1980-х годов; Национальный парк Маново-Гунда Сент-Флорис и Национальный парк и биосферный заповедник Баминги-Бангоран в Центральноафриканской Республике, где население сокращается; Заповедник Руманьяка Оругунду и Заповедник Ибанда в Танзании; и Национальный парк Мерчисон-Фолс в Уганда.[19]
- Хищник Лихтенштейна занесен в список наименьших проблем и встречается на охраняемых территориях, таких как Заповедник Селус и в дикой природе на юге и западе Танзании и Замбии.[72]
- В красный hartebeest внесен в список «Наименее вызывающий беспокойство». Это наиболее широко распространенный вид, численность которого увеличивается после возвращения в охраняемые и частные районы. Однако он вымер в Лесото с ХХ века.[19] Его население оценивается в более чем 130 000 человек (по данным на 2008 г.).[22] в основном в южной части Африки.[67] В Намибии самая большая популяция приходится на Национальный парк Этоша. Реинтродуцированное население процветает в Малолотский заповедник (Свазиленд ), за пределами его диапазона. Однако численность населения на юго-западе Ботсваны резко упала.[19]
- В Тора hartebeest указан как Находящихся под угрозой исчезновения; МСОП установил, что по состоянию на 2008 г. выжило менее 250 взрослых особей. Возможно, они вымерли в Судане из-за чрезмерной охоты и расширение сельского хозяйства, но все еще могут существовать в меньшем количестве в Эритрее и Эфиопии.[73] Поступали неподтвержденные сообщения о наблюдении местными жителями хартбеста Тора к юго-востоку от Национальный парк Диндер, откуда он исчез до 1960 года.[19]
- Житель Суэйна указан как Находящихся под угрозой исчезновения, и близок к тому, чтобы быть Находящихся под угрозой исчезновения. Общая численность популяции в 2008 году составляла менее 600, из которых взрослые особи насчитывали не более 250. Она ограничена четырьмя основными охраняемыми территориями: Заповедник дикой природы Сенкеле, Национальный парк Нечисар, Национальный парк Аваш и Национальный парк Mazie.[74] Зверобоям в Сенкеле приходится конкурировать с домашним скотом Люди оромо.[26] Исследование, проведенное в Национальном парке Нечисар в 2009 и 2010 годах, показало, что значительное увеличение поголовья скота оромо (на 49,9% и 56,5% в 2006 и 2010 годах соответственно), незаконная эксплуатация ресурсов и потеря среды обитания являются серьезными угрозами для хищных животных Суэйнов. населения там.[75]
- В западный hartebeest указан как Под угрозой.[76] Он был исключен из большей части своего ареала, включая юго-западные саванны и Национальный парк Букле-дю-Бауле в Мали; юго-западный Нигер; южный Сенегал; Гамбия; Кот-д'Ивуар; Буркина-Фасо. Небольшие группы населения выживают в Национальный парк Бафинг и площадь, ограниченная Бамако, Бугуни и Сикассо в Мали; Tamou Reserve в Нигере; Национальный парк Ниоколо-Коба в Сенегале; Национальный парк Комоэ в Кот-д'Ивуаре; Дифула лес и Ранчо Nazinga Game в Буркина-Фасо; Национальный парк Пенджари в Бенине; и Буба Нджида, Bénoué, и Фаро Национальные парки Камеруна.[19]
Отношения с людьми
Hartebeest популярны игра и трофей животные, поскольку они хорошо видны и, следовательно, на них легко охотиться.[38][63] Иллюстрированный, а также эпиграфический доказательства из Египет предполагает, что в Возраст верхнего палеолита Египтяне охотились на хищников и приручили их. Хартебист был важным источником мяса,[77] но его экономическое значение было ниже, чем у газелей и других пустынных видов.[48] Однако с начала эпохи неолита охота стала менее распространенной, и, следовательно, останки хищника этого периода в Египте, где он сейчас вымер, редки.[77]
В исследовании влияния места и пола на характеристики туши средний вес туши самцов красного хищника составлял 79,3 кг (175 фунтов), а самок - 56 кг (123 фунта). Мясо животных из региона Куа-Куа имело самый высокий липид содержание - 1,3 г (0,046 унции) на 100 г (3,5 унции) мяса. Незначительные различия были обнаружены в концентрациях отдельных жирные кислоты, аминокислоты, и минералы. В ходе исследования мясо хищника считалось полезным для здоровья, так как соотношение полиненасыщенный к насыщенные жирные кислоты составил 0,78, что немного больше рекомендованного 0,7.[78]
Рекомендации
- ^ а б c d е Группа специалистов МСОП по антилопам (2019). «Alcelaphus buselaphus (исправленная версия оценки 2016 г.)». Красный список видов, находящихся под угрозой исчезновения МСОП. 2019: e.T811A143160967. Дои:10.2305 / IUCN.UK.2019-1.RLTS.T811A143160967.en.
- ^ а б c d е Wilson, D.E .; Ридер, Д. М., ред. (2005). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Балтимор, США: Издательство Университета Джона Хопкинса. п. 674. ISBN 978-0-8018-8221-0. OCLC 62265494.CS1 maint: ref = harv (связь)
- ^ а б "Hartebeest". Словарь Merriam-Webster. Получено 24 января 2016.
- ^ а б Марес, М.А. (1999). Энциклопедия пустынь. Норман, США: Университет Оклахомы. п. 265. ISBN 978-0-8061-3146-7.
- ^ Ллевеллин, Э. К. (1936). «Влияние южноафриканского голландского языка или африкаанс на английский словарь». Влияние нижнего нидерландского языка на словарный запас английского языка. Лондон, Великобритания: Издательство Оксфордского университета. п. 163.
- ^ Скиннер, Дж. Д .; Чимимба, К. Т. (2005). Млекопитающие южноафриканского субрегиона (3-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. п. 649. ISBN 978-0-521-84418-5.
- ^ а б c d е ж грамм час Кингдон, Дж. (2013). Млекопитающие Африки. Лондон, Великобритания: Блумсбери. С. 510–22. ISBN 978-1-4081-2257-0.
- ^ «Конгони». Словарь Merriam-Webster. Получено 26 января 2016.
- ^ Суонк, У. Г. (1971). Африканская антилопа. Нью-Йорк, США: Winchester Press. п. 95. ISBN 978-0-87691-029-0.
- ^ а б c d Новак, Р. М. (1999). Млекопитающие Уокера (6-е изд.). Балтимор, США: Издательство Университета Джона Хопкинса. С. 1181–3. ISBN 978-0-8018-5789-8.
- ^ а б c Flagstad, Ø .; Syversten, P.O .; Stenseth, N.C .; Якобсен, К. С. (2001). «Изменение окружающей среды и темпы эволюции: филогеографические закономерности внутри комплекса Hartebeest, связанные с климатическими изменениями». Труды Королевского общества B: биологические науки. 268 (1468): 667–77. Дои:10.1098 / rspb.2000.1416. ЧВК 1088655. PMID 11321054.
- ^ Врба, Э. С. (1979). «Филогенетический анализ и классификация ископаемых и современных млекопитающих Alcelaphini: Bovidae». Биологический журнал Линнеевского общества. 11 (3): 207–28. Дои:10.1111 / j.1095-8312.1979.tb00035.x.
- ^ Джентри, А. В. (2012). «Эволюция и распространение африканских Bovidae». В Бубеник, Г. А .; Бубеник, А. Б. (ред.). Рога, вилорогие и рога: эволюция, морфология, физиология и социальное значение. Нью-Йорк, США: Springer. п. 216. ISBN 978-1-4613-8966-8.
- ^ Groves, C .; Грабб, П. (2011). Таксономия копытных. Балтимор, США: Издательство Университета Джона Хопкинса. п. 208. ISBN 978-1-4214-0093-8.
- ^ Matthee, C.A .; Робинсон, Т. Дж. (1999). «Филогения цитохрома b семейства Bovidae: разрешение в пределах Alcelaphini, Antilopini, Neotragini и Tragelaphini». Молекулярная филогенетика и эволюция. 12 (1): 31–46. Дои:10.1006 / mpev.1998.0573. PMID 10222159.
- ^ Arctander, P .; Johansen, C .; Coutellec-Vreto, M.A. (1999). «Филогеография трех близкородственных африканских быков (триба Alcelaphini)». Молекулярная биология и эволюция. 16 (12): 1724–39. Дои:10.1093 / oxfordjournals.molbev.a026085. PMID 10605114.
- ^ а б Flagstad, Ø .; Syvertsen, P.O .; Stenseth, N.C .; Стейси, Дж. Э .; Olsaker, I .; Røed, K. H .; Якобсен, К. С. (2000). "Генетическая изменчивость хищника Суэйна, находящейся под угрозой исчезновения антилопы Эфиопии". Биология сохранения. 14 (1): 254–64. Дои:10.1046 / j.1523-1739.2000.98339.x.
- ^ "Alcelaphus buselaphus". Интегрированная система таксономической информации. Получено 7 апреля 2016.
- ^ а б c d е ж грамм час я j k л м п Восток, р .; Группа специалистов по антилопам МСОП / SSC (1999). База данных африканских антилоп 1998. Гланд, Швейцария: Комиссия по выживанию видов МСОП. С. 186–93. ISBN 978-2-8317-0477-7.
- ^ а б Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus buselaphus". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 20 января 2013.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ Мэллон, Д. П .; Кингсвуд, С. С. (2001). Антилопы: Северная Африка, Ближний Восток и Азия.. Гланд, Швейцария: МСОП. п. 25. ISBN 978-2-8317-0594-1.
- ^ а б Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus caama". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 20 января 2013.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ а б Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus lelwel". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 23 января 2013.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ Рафферти, Дж. П. (2010). Grazers (1-е изд.). Нью-Йорк, США: Британские образовательные публикации. п. 121. ISBN 978-1-61530-465-3.
- ^ Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus lichtensteini". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 20 января 2013.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ а б Lewis, J. G .; Уилсон, Р. Т. (1977). "Бедственное положение жреца Суэйна". Орикс. 13 (5): 491–4. Дои:10.1017 / S0030605300014551.
- ^ а б Хилдьярд, А. (2001). Вымирающие дикие животные и растения мира. Нью-Йорк, США: Маршалл Кавендиш. С. 674–5. ISBN 978-0-7614-7199-8.
- ^ а б c d "Lelwel Hartebeest". Рекорды охоты на крупную дичь - Международная онлайн-книга рекордов Safari Club. Safari Club International. Получено 26 января 2016.
- ^ Robinson, T. J.; Моррис, Д. Дж .; Fairall, N. (1991). "Interspecific hybridisation in the Bovidae: Sterility of Alcelaphus buselaphus × Damaliscus dorcas F1 progeny". Биологическое сохранение. 58 (3): 345–56. Дои:10.1016/0006-3207(91)90100-N.
- ^ а б c d Castelló, J. R. (2016). Крупные рогатые животные мира: антилопы, газели, крупный рогатый скот, козы, овцы и родственники. Princeton, USA: Princeton University Press. pp. 537–9. ISBN 978-0-691-16717-6.
- ^ Augustine, D. J.; Veblen, K. E.; Goheen, J. R.; Riginos, C .; Young, T. P. (2011). "Pathways for positive cattle–wildlife interactions in semiarid rangelands" (PDF). In Georgiadis, N. J. (ed.). Conserving Wildlife in African Landscapes: Kenya's Ewaso Ecosystem. Вклад Смитсоновского института в зоологию. 632. Washington D.C., USA: Smithsonian Institution Scholarly Press. С. 55–71.
- ^ "Kenya Highland Hartebeest". Big Game Hunting Records – Safari Club International Online Record Book. Safari Club International. Получено 26 января 2016.
- ^ а б Ruxton, A. E.; Schwarz, E. (1929). "On hybrid hartebeests and on the distribution of the Alcelaphus buselaphus group". Труды Лондонского зоологического общества. 99 (3): 567–83. Дои:10.1111/j.1469-7998.1929.tb07706.x.
- ^ "Neumann Hartebeest". Big Game Hunting Records – Safari Club International Online Record Book. Safari Club International. Получено 26 января 2016.
- ^ Харрис, Дж .; Leaky, M. (2001). Лотагам: Рассвет человечества в Восточной Африке. New York, USA: Columbia University Press. п. 547. ISBN 978-0-231-11870-5.
- ^ Berger, L.R .; Hilton-Barber, B. (2004). Field Guide to the Cradle of Humankind: Sterkfontein, Swartkrans, Kromdraai & Environs World Heritage Site (2-е (перераб.) Изд.). Кейптаун, Южная Африка: Struik Publishers. п. 163. ISBN 978-1-77007-065-3.
- ^ Tsahar, E.; Izhaki, I.; Лев-Ядун, С .; Bar-Oz, G.; Hansen, D. M. (2009). "Distribution and extinction of ungulates during the Holocene of the southern Levant". PLOS ONE. 4 (4): 5316–28. Bibcode:2009PLoSO...4.5316T. Дои:10.1371/journal.pone.0005316. ЧВК 2670510. PMID 19401760.

- ^ а б c d е ж грамм час я j Kingdon, J. (1989). East African Mammals: An Atlas of Evolution in Africa. Volume 3, Part D: Bovids. Чикаго: Издательство Чикагского университета. ISBN 978-0-226-43725-5.
- ^ а б Macdonald, D. (1987). Энциклопедия млекопитающих. New York, USA: Facts on File. стр.564–71. ISBN 978-0-87196-871-5.
- ^ Capellini, I.; Gosling, L. M. (2007). "Habitat primary production and the evolution of body size within the hartebeest clade". Биологический журнал Линнеевского общества. 92 (3): 431–40. Дои:10.1111/j.1095-8312.2007.00883.x.
- ^ а б c "Western Hartebeest". Big Game Hunting Records – Safari Club International Online Record Book. Safari Club International. Получено 26 января 2016.
- ^ а б c "Cape or Red Hartebeest". Big Game Hunting Records – Safari Club International Online Record Book. Safari Club International. Получено 26 января 2016.
- ^ а б "Coke Hartebeest". Big Game Hunting Records – Safari Club International Online Record Book. Safari Club International. Получено 26 января 2016.
- ^ а б c "Lichtenstein Hartebeest". Big Game Hunting Records – Safari Club International Online Record Book. Safari Club International. Получено 26 января 2016.
- ^ а б c "Swayne Hartebeest". Big Game Hunting Records – Safari Club International Online Record Book. Safari Club International. Получено 26 января 2016.
- ^ а б c d е ж грамм час я j k л м Эстес, Р. Д. (2004). Руководство по поведению африканских млекопитающих: включая копытных, плотоядных и приматов (4-е изд.). Berkeley, USA: University of California Press. стр.133–42. ISBN 978-0-520-08085-0.
- ^ Firestone, M. (2009). Watching Wildlife: Southern Africa; South Africa, Namibia, Botswana, Zimbabwe, Malawi, Zambia (2-е изд.). Footscray, Australia: Lonely Planet. С. 228–9. ISBN 978-1-74104-210-8.
- ^ а б c Heckel, J. O. (2007). The present status of the hartebeest subspecies with special focus on north-east Africa and the Tora hartebeest (PDF) (Отчет). Ethiopian Wildlife Conservation Authority. С. 1–13. Архивировано из оригинал (PDF) 3 февраля 2016 г.. Получено 26 января 2016.
- ^ а б c d Capellini, I. (2007). "Dimorphism in the hartebeest". In Fairbairn, D. J.; Blanckenhorn, W. U.; Székely, T. (eds.). Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism. Лондон, Великобритания: Издательство Оксфордского университета. pp. 124–32. Дои:10.1093/acprof:oso/9780199208784.003.0014. ISBN 978-0-19-954558-2.
- ^ а б Capellini, I.; Gosling, L. M. (2006). "The evolution of fighting structures in hartebeest" (PDF). Исследования эволюционной экологии. 8: 997–1011.
- ^ Oboussier, H. (1970). "Information on Alcelaphini (Bovidae-Mammalia) with special reference to the brain and hypophysis. Results of research trips through Africa (1959–1967)". Gegenbaurs Morphologisches Jahrbuch. 114 (3): 393–435. PMID 5523305.
- ^ Schaller, G. B. (1976). Лев Серенгети: исследование отношений хищник-жертва. Чикаго, США: University of Chicago Press. стр.461–5. ISBN 978-0-226-73640-2.
- ^ Eltringham, S. K. (1979). The Ecology and Conservation of Large African Mammals (1-е изд.). Лондон, Великобритания: MacMillan. п. 177. ISBN 978-0-333-23580-5.
- ^ Verlinden, A. (1998). "Seasonal movement patterns of some ungulates in the Kalahari ecosystem of Botswana between 1990 and 1995". Африканский журнал экологии. 36 (2): 117–28. Дои:10.1046/j.1365-2028.1998.00112.x (inactive 2020-11-01).CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (связь)
- ^ Boomker, J.; Horak, I. G.; De Vos, V. (1986). "The helminth parasites of various artiodactylids from some South African nature reserves". Журнал ветеринарных исследований Onderstepoort. 53 (2): 93–102. PMID 3725333.
- ^ а б Howard, G. W. (1977). "Prevalence of nasal bots (Diptera: Oestridiae) in some Zambian hartebeest". Журнал болезней дикой природы. 13 (4): 400–4. Дои:10.7589/0090-3558-13.4.400. PMID 24228960. S2CID 27306683.
- ^ Pester, F. R. N.; Laurence, B. R. (2009). "The parasite load of some African game animals". Журнал зоологии. 174 (3): 397–406. Дои:10.1111/j.1469-7998.1974.tb03167.x.
- ^ Spitalska, E.; Riddell, M.; Heyne, H.; Sparagano, O.A. (2005). "Prevalence of theileriosis in red hartebeest (Alcelaphus buselaphus caama) in Namibia". Паразитологические исследования. 97 (1): 77–9. Дои:10.1007/s00436-005-1390-y. ISSN 1432-1955. PMID 15986252. S2CID 23721115.
- ^ Spinage, C. A. (2012). African Ecology: Benchmarks and Historical Perspectives. Берлин, Германия: Springer. п. 1176. ISBN 978-3-642-22872-8.
- ^ Belem, A. M. G.; Bakoné, É. U. (2009). "Parasites gastro-intestinaux d'antilopes et de buffles (Syncerus caffer brachyceros) du ranch de gibier de Nazinga au Burkina Faso" [Gastro-intestinal parasites of antelopes and buffaloes (Syncerus caffer brachyceros) from the Nazinga game ranch in Burkina Faso]. Biotechnologie, Agronomie, Société et Environnement (На французском). 13 (4): 493–8. ISSN 1370-6233.
- ^ Hoberg, E. P.; Abrams, A.; Pilitt, P. A. (2009). "Robustostrongylus aferensis ген. ноя et sp. ноя (Nematoda: Trichostrongyloidea) in kob (Кобус Коб) and hartebeest (Alcelaphus buselaphus jacksoni) (Artiodactyla) from sub-Saharan Africa, with further ruminations on the Ostertagiinae". Журнал паразитологии. 95 (3): 702–17. Дои:10.1645/ge-1859.1. PMID 19228080. S2CID 7641994. Получено 2018-04-29.
- ^ Le Roux, P. L. (1931). "На Longistrongylus meyeri ген. and sp. nov., a trichostrongyle parasitizing the Red Hartebeest Bubalis caama". Журнал гельминтологии. 9 (3): 141. Дои:10.1017/S0022149X00030376.
- ^ а б c d е ж "Hartebeest". Африканский фонд дикой природы. Получено 20 января 2013.
- ^ а б c Schuette, J. R.; Leslie, D. M.; Lochmiller, R. L.; Jenks, J. A. (1998). "Diets of hartebeest and Roan antelope in Burkina Faso: Support of the long-faced hypothesis". Журнал маммологии. 79 (2): 426–36. Дои:10.2307/1382973. JSTOR 1382973.
- ^ Murray, M. G. (1993). "Comparative nutrition of wildebeest, hartebeest and topi in the Serengeti". Африканский журнал экологии. 31 (2): 172–7. Дои:10.1111/j.1365-2028.1993.tb00530.x.
- ^ Casebeer, R. L.; Koss, G. G. (1970). "Food habits of wildebeest, zebra, hartebeest and cattle in Kenya Masailand". Африканский журнал экологии. 8 (1): 25–36. Дои:10.1111/j.1365-2028.1970.tb00827.x.
- ^ а б Миллс, G .; Hes, L. (1997). Полная книга южноафриканских млекопитающих. Кейптаун, Южная Африка: Struik Publishers. п. 255. ISBN 978-0-947430-55-9.
- ^ Yadav, P. R. (2004). Vanishing and Endangered Species. New Delhi, India: Discovery Publishing House. С. 139–40. ISBN 978-81-7141-776-6.
- ^ Mallon, D. P.; Kingswood, S. C. (2001). Антилопы: Северная Африка, Ближний Восток и Азия.. Гланд, Швейцария: МСОП. ISBN 978-2-8317-0594-1.
- ^ Harper, F. (1945). Extinct and Vanishing Mammals of the Old World. New York, USA: American Committee for International Wildlife Protection. pp. 642–8.
- ^ Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus cokii". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 23 февраля 2016.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus lichtensteinii". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 23 апреля 2013.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus tora". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 23 января 2013.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus swaynei". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 24 апреля 2013.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ Datiko, D.; Bekele, A. (2011). "Population status and human impact on the endangered Swayne's hartebeest (Alcelaphus buselaphus swaynei) in Nechisar Plains, Nechisar National Park, Ethiopia". Африканский журнал экологии. 49 (3): 311–9. Дои:10.1111/j.1365-2028.2011.01266.x.
- ^ Группа специалистов МСОП по антилопам (2008 г.). "Alcelaphus buselaphus major". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 23 января 2013.CS1 maint: использует параметр авторов (связь) CS1 maint: ref = harv (связь)
- ^ а б Van Neer, W.; Linseele, V.; Friedman, R. F. (2004). "Animal burials and food offerings at the elite cemetery HK6 of Hierakonpolis". In Hendrickx, S.; Фридман, Р. Ciałowicz, K.; Chłodnicki, M. (eds.). Egypt at its Origins: Studies in Memory of Barbara Adams. Orientalia Lovaniensia Analecta. 138. Leuven, Belgium: Peeters Publishers. п. 111. ISBN 978-90-429-1469-8.
- ^ Hoffman, L. C.; Smit, K.; Muller, N. (2010). "Chemical characteristics of red hartebeest (Alcelaphus buselaphus caama) meat". South African Journal of Animal Science. 40 (3): 221–8. Дои:10.4314/sajas.v40i3.6.
внешняя ссылка
 СМИ, связанные с Alcelaphus buselaphus в Wikimedia Commons
СМИ, связанные с Alcelaphus buselaphus в Wikimedia Commons Данные, относящиеся к Alcelaphus buselaphus в Wikispecies
Данные, относящиеся к Alcelaphus buselaphus в Wikispecies
| Авторитетный контроль |
|---|