Вирус эшерихии Т4 - Escherichia virus T4

| Вирус эшерихии Т4 | |
|---|---|
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Дуплоднавирия |
| Королевство: | Heunggongvirae |
| Тип: | Уровирикота |
| Учебный класс: | Caudoviricetes |
| Заказ: | Caudovirales |
| Семья: | Myoviridae |
| Род: | Текватровирус |
| Разновидность: | Вирус эшерихии Т4 |
| Штаммы[1] | |
| Синонимы[2] | |
Энтеробактерии фаг Т4 | |
Вирус эшерихии Т4 это разновидность бактериофаги это заразить кишечная палочка бактерии. Это двухцепочечный ДНК-вирус из подсемейства Tevenvirinae из семьи Myoviridae. Т4 способен пройти только литический жизненный цикл а не лизогенный жизненный цикл. Вид был ранее назван Т-четный бактериофаг, название, которое также включает, среди других штаммов (или изолятов), Энтеробактерии фаг Т2, Энтеробактерии фаг Т4 и Энтеробактерии фаг Т6.
Бактериофаг означает «поедать бактерии», а фаги, как известно, являются облигатными внутриклеточными паразиты которые воспроизводятся в клетке-хозяине и высвобождаются, когда хозяин уничтожается лизис. Полный геном последовательность фага Т4 содержит 168 903 пар оснований и кодирует около 300 генные продукты.[3] Эти вирулентные вирусы являются одними из самых крупных и сложных известных вирусов и одними из наиболее изученных. модельные организмы. Они сыграли ключевую роль в развитии вирусология и молекулярная биология.[4][5]
Использование в исследованиях
Начиная с 1940-х годов и по сей день, Т-четные фаги считаются наиболее изученными модельными организмами. Модельные организмы обычно требуются простые, всего пять гены. Тем не менее, Т-четные фаги на самом деле являются одними из самых больших и сложных вирус, в котором эти фаги генетическая информация состоит из около 300 гены. Совпадая с их сложностью, Т-четные вирусы не обладали невообразимой особенностью - наличием необычного основания. гидроксиметилцитозин (HMC) вместо основания нуклеиновой кислоты цитозин. В дополнение к этому, остатки HMC на Т-четном фаге глюкозилируются по определенной схеме. Эта уникальная особенность позволила сформировать новые ферменты, которые никогда не существовали в T-четных инфицированных клетках или в любых других клетках, а модификация ДНК T-четного фага обеспечила фундаментальный прогресс в популярный и молекулярный уровни. Другой уникальной особенностью вируса T-even является его регулируемая экспрессия генов.[6]
Геном и структура
Двухцепочечный вирус Т4 ДНК геном около 169 kbp длинный[7] и кодирует 289 белки. Геном Т4 окончательно избыточен. При репликации ДНК образуются длинные конкатемеры, имеющие несколько длин геномов, возможно, по механизму репликации катящегося круга.[8] При упаковке конкатемер разрезается в неспецифических положениях одинаковой длины, что приводит к нескольким геномам, которые представляют круговые перестановки оригинала.[9] Геном Т4 несет эукариот -подобно интрон последовательности.
Перевод
В Последовательность Шайна-Далгарно GAGG доминирует в ранних генах вируса T4, тогда как последовательность GGAG является мишенью для T4 эндонуклеаза RegB, который инициирует раннюю деградацию мРНК.[10]
Структура вирусных частиц

Т4 - относительно большой вирус, примерно 90%. нм шириной и длиной 200 нм (большинство вирусов имеют длину от 25 до 200 нм). Геном ДНК хранится в икосаэдр голова, также известная как капсид.[11] Хвост Т4 полый, так что он может проходить нуклеиновая кислота в клетку он заражает после прикрепления. Фаги Myoviridae, такие как T4, имеют сложные структуры сократительного хвоста с большим количеством белков, участвующих в сборке и функционировании хвоста.[12] Хвостовые волокна также важны для распознавания рецепторов на поверхности клетки-хозяина, поэтому они определяют, находится ли бактерия в пределах диапазона хозяина вируса.[13]
Недавно была описана структура 6-мегадальтонной базовой пластины Т4, которая включает 127 полипептидных цепей 13 различных белков (генные продукты 5, 5.4, 6, 7, 8, 9, 10, 11, 12, 25, 27, 48 и 53). в атомарных деталях. Также была создана атомная модель проксимальной области хвостовой трубки, образованной gp54 и белком основной трубки gp19. Белок gp29 с рулеткой присутствует в комплексах базовая пластинка-хвостовая трубка, но его нельзя смоделировать.[14]
Процесс заражения
Вирус Т4 инициирует кишечная палочка инфекционное заболевание путем привязки OmpC порин белки и липополисахарид (LPS) на поверхности Кишечная палочка клетки с длинными хвостовыми волокнами (LTF).[15][16] Сигнал распознавания отправляется через LTF на опорную плиту. Это распутывает короткие хвостовые волокна (STF), которые необратимо связываются с Кишечная палочка клеточная поверхность. Опорная плита меняет форму, и оболочка хвоста сжимается, в результате чего GP5 на конце хвостовой трубы пробивает внешнюю мембрана ячейки.[17] В лизоцим домен GP5 активируется и разрушает периплазматический пептидогликан слой. Оставшаяся часть мембраны разрушается, и тогда ДНК из головы вируса может пройти через хвостовую трубку и попасть в Кишечная палочка клетка.
Размножение
В литический жизненный цикл (от попадания в бактерию до ее уничтожения) занимает около 30 минут (при 37 ° C). Вирулентные бактериофаги размножаются в своем бактериальном хозяине сразу после проникновения. После того, как количество фагов-потомков достигнет определенного количества, они заставят хозяина лизироваться или разрушаться, поэтому они будут высвобождаться и заражать новые клетки-хозяева.[18] Процесс лизиса и высвобождения хозяина называется литический цикл. Литический цикл - это цикл размножения вируса, который включает разрушение инфицированной клетки и ее мембраны. В этом цикле участвует вирус, который заставляет клетку-хозяина и ее механизмы воспроизводиться. Следовательно, вирус должен пройти 5 стадий, чтобы воспроизвести и заразить клетку-хозяина:
- Адсорбция и проникновение (начинается сразу)
- Арест хозяина экспрессия гена (начиная немедленно)
- Фермент синтез (запуск через 5 минут)
- Репликация ДНК (начало через 10 минут)
- Формирование новых вирус частицы (начиная через 12 минут)
После завершения жизненного цикла клетка-хозяин разрывается и выбрасывает недавно созданные вирусы в окружающую среду, разрушая клетку-хозяин. T4 имеет размер взрыва приблизительно 100-150 вирусных частиц на инфицированного хозяина.
Бензер (1955 - 1959) разработал систему для изучения тонкой структуры гена с использованием мутантов бактериофага Т4, дефектных по rIIA и rIIB гены.[19][20][21] Использованные методы были дополнение тесты и кресты для обнаружения рекомбинация, особенно между делеционными мутациями. Эти генетические эксперименты привели к обнаружению уникального линейного порядка мутационных сайтов в генах. Этот результат явился убедительным доказательством ключевой идеи о том, что ген имеет линейную структуру, эквивалентную длине ДНК, со многими сайтами, которые могут независимо мутировать.
Адсорбция и проникновение

Как и все другие вирусы, T-четные фаги не просто случайно прикрепляются к поверхности своего хозяина; вместо этого они "ищут" и привязываются к рецепторы, специфический белок структуры, обнаруженные на поверхности хозяина. Эти рецепторы различаются в зависимости от фага; тейхоевая кислота, белки клеточной стенки и липополисахариды, жгутики, и пили все они могут служить рецепторами для связывания фага. Чтобы Т-четный фаг заразил своего хозяина и начал свой жизненный цикл, он должен войти в первый процесс инфекционное заболевание, адсорбция фага в бактериальную клетку. Адсорбция - это ценностная характеристика пары фаг-хозяин, и адсорбция фага на поверхности клетки-хозяина проиллюстрирована как двухэтапный процесс: обратимый и необратимый. Он включает структуру хвоста фага, которая начинается, когда волокна хвоста фага помогают связать фаг с соответствующим рецептором его хозяина. Этот процесс обратимый. Один или несколько компонентов базовой пластины опосредуют необратимый процесс связывания фага с бактерией.
Проникновение также является ценностной характеристикой фага-хозяина. инфекционное заболевание что включает в себя инъекцию фагов генетический материал внутри бактерия. Проникновение нуклеиновая кислота происходит после фазы необратимой адсорбции. Механизмы проникновения нуклеиновой кислоты фага специфичны для каждого фага. Этот механизм проникновения может включать электрохимический мембранный потенциал, АТФ молекулы, ферментативное расщепление пептидогликан слой, или все три из этих факторов могут быть жизненно важными для проникновения нуклеиновой кислоты внутрь бактериальной клетки. Исследования были проведены на Бактериофаг Т2 (Т4-подобный фаг) механизм проникновения, и было показано, что хвост фага не проникает внутрь стенки бактериальной клетки, а проникновение этого фага включает электрохимические мембранный потенциал на внутренней мембране.[22][23]
Репликация и упаковка
Геном вируса Т4 синтезируется в клетке-хозяине с использованием репликации по катящемуся кругу.[8] Время, необходимое для репликации ДНК в живой клетке, измеряли как скорость удлинения ДНК вируса Т4 в инфицированной вирусом E. coli.[24] В период экспоненциального увеличения ДНК при 37 ° C скорость составила 749 нуклеотидов в секунду. Частота мутаций на пару оснований на репликацию во время синтеза ДНК вируса Т4 составляет 1,7 на 10−8,[25] высокоточный механизм копирования ДНК, всего 1 ошибка на 300 копий. Вирус также кодирует уникальные Ремонт ДНК механизмы.[26] Головка фага Т4 пуста собирается вокруг каркасного белка, который позже разрушается. Следовательно, ДНК должна попасть в головку через крошечную пору, что достигается за счет взаимодействия гексамера gp17 с ДНК, которая также служит двигателем и нуклеазой. Было обнаружено, что двигатель упаковки ДНК Т4 загружает ДНК в вирусные капсиды со скоростью до 2000 пар оснований в секунду. Потребляемая мощность, если ее увеличить в размерах, будет эквивалентна мощности среднего автомобильного двигателя.[27]
Трансдукция происходит в литическом цикле, когда единицы бактериальной ДНК упаковываются в фаговый капсид.[28]
Релиз
Заключительный этап размножения и размножения вирусов определяется высвобождением вирионов из клетки-хозяина. Высвобождение вирионов происходит после разрыва плазматической мембраны бактерий. Вирусы без оболочки лизируют клетку-хозяин, которая характеризуется вирусными белками, атакующими пептидогликан или мембрану. Лизис бактерий происходит, когда капсиды внутри клетки высвобождают фермент лизоцим, который разрушает клеточную стенку. Освободившиеся бактериофаги инфицируют другие клетки, и цикл размножения вируса повторяется внутри этих клеток.
Повторная активация множественности
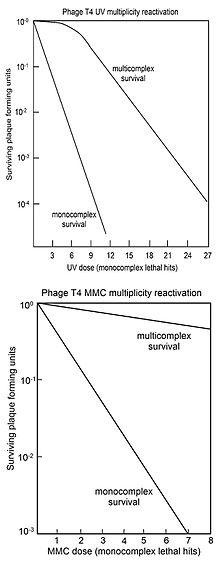
Реактивация множественности (MR) - это процесс, при котором два или более вирусных генома, каждый из которых содержит инактивирующее повреждение генома, могут взаимодействовать в инфицированной клетке с образованием жизнеспособного вирусного генома. Сальвадор Луриа, изучая в 1946 г. облученный УФ-излучением вирус Т4, открыл MR и предположил, что наблюдаемая реактивация поврежденного вируса происходит по механизму рекомбинации (см. Ссылки.[29][30][31]) Это предшествовало подтверждению ДНК как генетического материала в 1952 г. в родственном вирусе Т2 Эксперимент Херши – Чейза.[32]
Как вспоминал Лурия (1984,[33] стр. 97) открытие реактивации облученного вируса (именуемого «повторная активация множественности ") сразу же начал бурную деятельность по изучению ремонта радиационных повреждений в первые фаговая группа (обзор Bernstein[26] в 1981 г.). Позже выяснилось, что восстановление поврежденного вируса взаимной помощью, которое обнаружил Лурия, было лишь частным случаем восстановления ДНК. Клетки всех типов, не только бактерии и их вирусы, но и все изученные организмы, включая человека, теперь, как известно, обладают сложными биохимическими процессами восстановления повреждений ДНК (см. Ремонт ДНК ). В настоящее время признается, что процессы репарации ДНК играют важную роль в защите от старение, рак, и бесплодие.
MR обычно представляет собой «кривые выживаемости», где выживаемость бляшкообразующей способности многократно инфицированных клеток (мультикомплексов) отображается в зависимости от дозы агента, повреждающего геном. Для сравнения, выживаемость вирусного бляшкообразования отдельно инфицированных клеток (монокомплексов) также наносится на график в зависимости от дозы агента, повреждающего геном. На верхнем рисунке показаны кривые выживаемости мультикомплексов и монокомплексов вируса Т4 при увеличении дозы УФ-излучения. Поскольку выживаемость нанесена на логарифмическую шкалу, ясно, что выживаемость мультикомплексов превышает выживаемость монокомплексов в очень большие факторы (в зависимости от дозы). Кривая УФ-инактивации для мультикомплексов имеет начальное плечо. Другими агентами, повреждающими ДНК вируса Т4 с плечом в их мультикомплексных кривых выживания, являются рентгеновские лучи.[34][35] и этилметансульфонат (EMS).[26] Присутствие плеча было интерпретировано как означающее, что используются два рекомбинационных процесса.[36] Первый восстанавливает ДНК с высокой эффективностью (в «плече»), но насыщается своей способностью по мере увеличения повреждений; второй путь действует на всех уровнях повреждения. Выживший вирус Т4, высвобожденный из мультикомплексов, не показывает увеличения мутация, что указывает на то, что MR вируса, облученного УФ-излучением, является точным процессом.[36]
На нижнем рисунке показаны кривые выживаемости при инактивации вируса Т4 повреждающим ДНК агентом. митомицин С (MMC). В этом случае кривая выживаемости для мультикомплексов не имеет начального плеча, что позволяет предположить, что активен только второй процесс рекомбинационной репарации, описанный выше. На эффективность восстановления с помощью этого процесса указывает наблюдение, что доза MMC, которая позволяет выжить только 1 из 1000 монокомплексов, позволяет выжить примерно 70% мультикомплексов. Аналогичные мультикомплексные кривые выживаемости (без плеч) были получены и для повреждающих ДНК агентов. P32 разлагаться, псорален плюс ближнее УФ-облучение (ПУВА), N-метил-N'-нитро-N-нитрозогуанидин (MNNG), метилметансульфонат (MMS) и азотистая кислота.[26]
Некоторые из генов, необходимых для MR в вирусе T4, оказались ортологи для генов, необходимых для рекомбинации в прокариоты, эукариоты и археи. Это включает, например, ген Т4. uvsX[37] который определяет белок, имеющий трехмерную структурную гомологию с RecA из кишечная палочка и гомологичный белок RAD51 в эукариоты и RadA в археи. Было высказано предположение, что эффективная и точная рекомбинационная репарация повреждений ДНК во время MR может быть аналогична процессу рекомбинационной репарации, который происходит во время мейоз в эукариоты.[38]
История
Бактериофаги были впервые обнаружены английским ученым. Фредерик Творт в 1915 г. и Феликс д'Эрелль в 1917 г. В конце 1930-х гг. Т.Л. Ракитен предложил смесь неочищенных сточных вод или лизат из Кишечная палочка заражены сырой канализацией двух исследователей Милислав Демерец и Уго Фано. Эти два исследователя изолировали Т3, Т4, Т5 и Т6 от Кишечная палочка. Также в 1932 году исследователь Дж. Бронфенбреннер изучал и работал над фагом Т2, на котором фаг Т2 был выделен из вируса.[39] Эта изоляция была сделана из фекалий, а не из канализации. Во всяком случае, Дельбрук участвовал в открытии Т-четных фагов. Его часть заключалась в присвоении бактериофагов типу 1 (T1), типу 2 (T2), типу 3 (T3) и т. Д.
Конкретное время и место выделения вируса Т4 остается неясным, хотя, вероятно, они были обнаружены в сточных водах или фекалиях. T4 и подобные вирусы были описаны в статье Томас Ф. Андерсон, Макс Дельбрюк, и Милислав Демерец в ноябре 1944 г.[40]
В фаговая группа была неформальной сетью биологов, сосредоточенной на Макс Дельбрюк который провел фундаментальные исследования, главным образом, на бактериофаге Т4, и внес значительный вклад в микробная генетика и истоки молекулярной биологии в середине 20 века. В течение 1962-1964 гг. Эта группа исследователей фага Т4 предоставила возможность изучить функцию практически всех генов, которые необходимы для роста фага в лабораторных условиях.[41][42] Этим исследованиям способствовало открытие двух классов условно-летальные мутанты. Один класс таких мутантов известен как янтарные мутанты.[43] Другой класс условно-летальных мутантов называется термочувствительные мутанты[44] Исследования этих двух классов мутантов привели к глубокому пониманию многих фундаментальных биологических проблем. Таким образом было получено понимание функций и взаимодействий белков, используемых в механизме Репликация ДНК, ремонт и рекомбинация и как вирусы собираются из компонентов белка и нуклеиновых кислот (молекулярные морфогенез ). Кроме того, роль завершающие кодоны цепи было выяснено. В одном заслуживающем внимания исследовании использовались мутанты янтаря, дефектные по гену, кодирующему главный головной белок фага Т4.[45] Этот эксперимент предоставил убедительные доказательства широко распространенной, но до 1964 года еще не доказанной «гипотезы последовательности» о том, что аминокислотная последовательность белка определяется нуклеотидная последовательность из ген определение белка. Таким образом, это исследование продемонстрировало коллинеарность гена с кодируемым им белком.
Номер Нобелевская премия победители работали с вирусом Т4 или Т4-подобными вирусами, включая Макс Дельбрюк, Сальвадор Лурия, Альфред Херши, Джеймс Д. Уотсон, и Фрэнсис Крик. Другие важные ученые, работавшие с вирусом T4, включают: Майкл Россманн, Сеймур Бензер, Брюс Альбертс, Гизела Мосиг,[46] Ричард Ленски, и Джеймс Булл.
Смотрите также
Рекомендации
- ^ "Девятый отчет ICTV (2011 г.) Myoviridae". Международный комитет по таксономии вирусов (ICTV). Получено 26 декабря 2018.
- ^ "История таксономии ICTV: Вирус эшерихии Т4". Международный комитет по таксономии вирусов (ICTV). Получено 26 декабря 2018.
Caudovirales > Myoviridae > Tevenvirinae > T4virus > Вирус эшерихии Т4
- ^ Миллер Э.С., Куттер Э., Мосиг Г., Арисака Ф., Кунисава Т., Рюгер В. Геном бактериофага Т4. Microbiol Mol Biol Rev.2003 Mar; 67 (1): 86-156, содержание. DOI: 10.1128 / mmbr.67.1.86-156.2003. PMID: 12626685; PMCID: PMC150520.
- ^ Норкин, Леонард С. (2010). Вирусология, молекулярная биология и патогенез. Вашингтон: Американское общество микробиологии. п. 725. ISBN 978-1-55581-453-3.
- ^ Прескотт, Харли и Кляйн (2008). Микробиология (седьмое изд.). Макгроу Хилл. п. 1078. ISBN 978-007-126727-4.CS1 maint: несколько имен: список авторов (связь)
- ^ Норкин, Леонард С. (2010). Вирусология, молекулярная биология и патогенез. Вашингтон: Американское общество микробиологии. п. 725. ISBN 978-1-55581-453-3.
- ^ Миллер, ES; Куттер, Э; Мосиг, G; Arisaka, F; Кунисава, Т; Рюгер, W (март 2003 г.). «Геном бактериофага Т4». Обзоры микробиологии и молекулярной биологии. 67 (1): 86–156, содержание. Дои:10.1128 / MMBR.67.1.86-156.2003. ЧВК 150520. PMID 12626685.
- ^ а б Бернштейн Х., Бернштейн С. Круглые и разветвленные кольцевые конкатенации как возможные промежуточные продукты в репликации ДНК бактериофага Т4. J Mol Biol. 5 июля 1973 г .; 77 (3): 355-61. DOI: 10.1016 / 0022-2836 (73) 90443-9. PMID: 4580243.
- ^ Мэдиган М., Мартинко Дж., Ред. (2006). Биология микроорганизмов Брока (11-е изд.). Прентис Холл. ISBN 978-0-13-144329-7.
- ^ Малыс Н (2012). «Последовательность Шайна-Далгарно бактериофага T4: GAGG преобладает в ранних генах». Отчеты по молекулярной биологии. 39 (1): 33–9. Дои:10.1007 / s11033-011-0707-4. PMID 21533668.
- ^ Прескотт, Харли и Кляйн (2008). Микробиология (седьмое изд.). Макгроу-Хилл. ISBN 978-007-126727-4.
- ^ Петр Г. Лейман, Фумио Арисака, Марк Дж. Ван Раай, Виктор А. Костюченко, Анастасия А. Аксюк, Шуджи Канамару и Майкл Г. Россманн. Морфогенез хвоста и волокон хвоста Т4, Журнал вирусологии, том 7, номер статьи: 355 (2010) https://virologyj.biomedcentral.com/articles/10.1186/1743-422X-7-355
- ^ Ackermann, H.-W .; Криш, Х. М. (6 апреля 2014 г.). «Каталог бактериофагов Т4-типа». Архив вирусологии. 142 (12): 2329–2345. Дои:10.1007 / s007050050246. PMID 9672598.
- ^ Тейлор, Николас М. И .; Прохоров, Николай С .; Герреро-Феррейра, Рикардо К.; Шнейдер, Михаил М .; Браунинг, Кристофер; Goldie, Kenneth N .; Штальберг, Хеннинг; Лейман, Петр Г. (2016). «Структура T4 опорной плиты и ее функции в инициировании сжатия оболочки». Природа. 533 (7603): 346–352. Bibcode:2016Натура.533..346Т. Дои:10.1038 / природа17971. PMID 27193680.
- ^ Ю, Ф .; Мидзусима, С. (1982). «Роль липополисахарида и белка внешней мембраны OmpC Escherichia coli K-12 в рецепторной функции бактериофага T4». Журнал бактериологии. 151 (2): 718–722. Дои:10.1128 / JB.151.2.718-722.1982. ЧВК 220313. PMID 7047495.
- ^ Furukawa, H .; Мидзусима, С. (1982). «Роль компонентов клеточной поверхности Escherichia coli K-12 в инфекции бактериофагом T4: взаимодействие ядра хвоста с фосфолипидами». Журнал бактериологии. 150 (2): 916–924. Дои:10.1128 / JB.150.2.916-924.1982. ЧВК 216445. PMID 7040345.
- ^ Maghsoodi, A .; Chatterjee, A .; Andricioaei, I .; Перкинс, Северная Каролина (25 ноября 2019 г.). «Как работает оборудование для инъекции фага Т4, включая энергию, силы и динамический путь». Труды Национальной академии наук. Труды Национальной академии наук. 116 (50): 25097–25105. Дои:10.1073 / pnas.1909298116. ISSN 0027-8424.
- ^ Шервуд, Линда (2011). Микробиология Прескотта (восьмое изд.). Макгроу-Хилл.
- ^ Бензер С. «Приключения в регионе rII» в книге «Фаг и происхождение молекулярной биологии» (2007) Под редакцией Джона Кэрнса, Гюнтера С. Стента и Джеймса Д. Уотсона, Лаборатория количественной биологии Колд-Спринг-Харбор, Колд-Спринг-Харбор, Лонг. Остров, Нью-Йорк ISBN 978-0879698003
- ^ Бензер С. Тонкая структура генетической области бактериофага. Proc Natl Acad Sci U S. A. 1955, 15 июня; 41 (6): 344-54. PMID 16589677
- ^ Бензер С. К топологии тонкой генетической структуры. Proc Natl Acad Sci U S. A. 1959 ноябрь; 45 (11): 1607-20. PMID 16590553
- ^ Норкин, Леонард С. (2010). Вирусология, молекулярная биология и патогенез. Вашингтон: Американское общество микробиологии. п. 31. ISBN 978-1-55581-453-3.
- ^ Прескотт, Харли и Кляйн (2008). Микробиология (седьмое изд.). Макгроу Хилл. п. 427. ISBN 978-007-126727-4.CS1 maint: несколько имен: список авторов (связь)
- ^ Маккарти Д., Миннер С., Бернштейн Х, Бернштейн С. (1976). «Скорость удлинения ДНК и распределение точек роста фага Т4 дикого типа и янтарного мутанта с задержкой ДНК». Дж Мол Биол. 106 (4): 963–81. Дои:10.1016/0022-2836(76)90346-6. PMID 789903.
- ^ Дрейк JW (1970) Молекулярные основы мутации. Холден-Дэй, Сан-Франциско ISBN 0816224501 ISBN 978-0816224500
- ^ а б c d Бернштейн С. "Ремонт дезоксирибонуклеиновой кислоты в бактериофаге". Microbiol Rev.1981 Mar; 45 (1): 72-98. Рассмотрение. PMID 6261109
- ^ Рао, Венигалла Б; Блэк, Линдси В. (1 января 2010 г.). «Строение и сборка головки бактериофага Т4». Журнал вирусологии. 7 (1): 356. Дои:10.1186 / 1743-422X-7-356. ЧВК 3012670. PMID 21129201.
- ^ Леонард С., Норкин (2010). Вирусология, молекулярная биология и патогенез. Вашингтон: Американское общество микробиологии.
- ^ Лурия С.Е. (1947). «Реактивация облученного бактериофага путем переноса самовоспроизводящихся единиц». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 33 (9): 253–64. Bibcode:1947ПНАС ... 33..253Л. Дои:10.1073 / pnas.33.9.253. ЧВК 1079044. PMID 16588748.
- ^ ЛУРИА СЕ, ДУЛЬБЕККО Р. (1948). «Летальные мутации и инактивация индивидуальных генетических детерминант бактериофага». Генетика. 33 (6): 618. PMID 18100306.
- ^ Лурия С.Е., Дульбекко Р. (1949). «Генетические рекомбинации, ведущие к производству активного бактериофага из частиц бактериофага, инактивированных ультрафиолетом». Генетика. 34 (2): 93–125. ЧВК 1209443. PMID 17247312.
- ^ ГЕРШИ Э.Д., ЧАС М (1952). «Независимые функции вирусного белка и нуклеиновой кислоты в росте бактериофага». J. Gen. Physiol. 36 (1): 39–56. Дои:10.1085 / jgp.36.1.39. ЧВК 2147348. PMID 12981234.
- ^ Сальвадор Э. Лурия. Игровой автомат, сломанная пробирка: автобиография. Harper & Row, Нью-Йорк: 1984. Стр. 228. ISBN 0-06-015260-5 (США и Канада)
- ^ Уотсон Дж. Д. (1952). «Свойства рентгеновского инактивированного бактериофага». J. Bacteriol. 63 (4): 473–85. Дои:10.1128 / JB.63.4.473-485.1952. ЧВК 169298. PMID 14938320.
- ^ ВРЕД W (1958). «Реактивация множественности, спасение маркеров и генетическая рекомбинация в фаге Т4 после инактивации рентгеновскими лучами». Вирусология. 5 (2): 337–61. Дои:10.1016/0042-6822(58)90027-8. PMID 13544109.
- ^ а б Ярош Д.Б. (1978). «УФ-индуцированная мутация бактериофага Т4». Дж. Вирол. 26 (2): 265–71. Дои:10.1128 / JVI.26.2.265-271.1978. ЧВК 354064. PMID 660716.
- ^ История Р.М., Епископ Д.К., Клекнер Н., Стейтц Т.А. (1993). «Структурная связь бактериальных белков RecA с белками рекомбинации из бактериофага Т4 и дрожжей». Наука. 259 (5103): 1892–6. Bibcode:1993Научный ... 259.1892S. Дои:10.1126 / science.8456313. PMID 8456313.
- ^ Бернштейн C (1979). «Почему дети маленькие? Мейоз может предотвратить старение зародышевой линии». Перспектива. Биол. Med. 22 (4): 539–44. Дои:10.1353 / pbm.1979.0041. PMID 573881.
- ^ Уилли, Джоанна. Микробиология Прескотта (седьмое изд.). Макгроу-Хилл.
- ^ Абедон, СТ (июнь 2000 г.). "Неясное происхождение Белоснежки и ее карликов-даже". Генетика. 155 (2): 481–6. ЧВК 1461100. PMID 10835374.
- ^ Эдгар Р.С. Условная летальность: в книге «Фаг и происхождение молекулярной биологии» (2007) Под редакцией Джона Кэрнса, Гюнтера С. Стента и Джеймса Д. Уотсона, Лаборатория количественной биологии Колд-Спринг-Харбор, Колд-Спринг-Харбор, Лонг-Айленд, Нью-Йорк ISBN 978-0879698003
- ^ Эдгар Б. (2004). «Геном бактериофага Т4: археологические раскопки». Генетика 168 (2): 575–82. PMC 1448817. PMID 15514035.
- ^ Эпштейн, Р.Х., Болле, К.М. Стейнберг, Э., Келленбергер, Э., Бой де ла Тур и др., 1964. Физиологические исследования условных летальных мутантов бактериофага T4D. Холодный источник Харб. Symp. Quant. Биол. 28: 375-394.
- ^ Эдгар Р.С., Лиелузис I. Термочувствительные мутанты бактериофага T4D: их выделение и характеристика. Генетика. 1964 апр; 49: 649-62. PMID 14156925
- ^ Сарабхай А.С., Стреттон А.О., Бреннер С., Болле А. Коллинеарность гена с полипептидной цепью. Природа. 1964, 4 января; 201: 13-7. PMID 14085558
- ^ Носал, НГ; Франклин, JL; Куттер, Э; Дрейк, JW (ноябрь 2004 г.). «Анекдотические, исторические и критические комментарии по генетике. Гизела Мосиг». Генетика. 168 (3): 1097–104. ЧВК 1448779. PMID 15579671.
дальнейшее чтение
- Leiman P.G .; Kanamaru S; Месянжинов В.В .; Arisaka F .; Россманн М.Г. (2003). «Структура и морфогенез бактериофага Т4». Клеточные и молекулярные науки о жизни. 60 (11): 2356–2370. Дои:10.1007 / s00018-003-3072-1. PMID 14625682.
- Карам, Дж., Петров, В., Нолан, Дж., Чин, Д., Шатли, К., Криш, Х., Летаров, А. Проект генома Т4-подобных фагов. https://web.archive.org/web/20070523215704/http://phage.bioc.tulane.edu/. (Депозитарий полных геномных последовательностей Т4-подобных фагов)
- Мосиг Г. и Ф. Эйзерлинг. 2006. T4 и родственные фаги: структура и развитие, Р. Календар и С. Т. Абедон (ред.), Бактериофаги. Издательство Оксфордского университета, Оксфорд. (Обзор биологии фага Т4) ISBN 0-19-514850-9
- Filee J. Tetart F .; Suttle C.A .; Krisch H.M. (2005). «Морские бактериофаги типа T4, вездесущий компонент темной материи биосферы». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 102 (35): 12471–6. Bibcode:2005PNAS..10212471F. Дои:10.1073 / pnas.0503404102. ЧВК 1194919. PMID 16116082. (Индикация распространенности и Т4-подобных фагов в дикой природе)
- Chibani-Chennoufi S .; Canchaya C .; Bruttin A .; Брюссов Х. (2004). «Сравнительная геномика Т4-подобного кишечная палочка фаг JS98: значение для эволюции фагов Т4 ». J. Bacteriol. 186 (24): 8276–86. Дои:10.1128 / JB.186.24.8276-8286.2004. ЧВК 532421. PMID 15576776. (Характеристика Т4-подобного фага)
- Desplats C, Krisch HM (май 2003 г.). «Разнообразие и эволюция бактериофагов Т4-типа». Res. Микробиол. 154 (4): 259–67. Дои:10.1016 / S0923-2508 (03) 00069-X. PMID 12798230.
- Miller, E.S .; Kutter E .; Mosig G .; Arisaka F .; Кунисава Т .; Ругер В. (2003). «Геном бактериофага Т4». Microbiol. Мол. Биол. Rev. 67 (1): 86–156. Дои:10.1128 / MMBR.67.1.86-156.2003. ЧВК 150520. PMID 12626685. (Обзор фага Т4 с точки зрения его генома)
- Desplats C .; Dez C .; Tetart F .; Eleaume H .; Krisch H.M. (2002). «Снимок генома псевдо-Т-четного бактериофага RB49». J. Bacteriol. 184 (10): 2789–2804. Дои:10.1128 / JB.184.10.2789-2804.2002. ЧВК 135041. PMID 11976309. (Обзор генома RB49, T4-подобного фага)
- Малис Н., Чанг Д.Й., Бауманн Р.Г., Се Д., Блэк Л.В. (2002). «Библиотека рандомизированного пептидного дисплея SOC и HOC бактериофага T4: обнаружение и анализ взаимодействия фага T4-терминазы (gp17) и фактора поздней сигмы (gp55)». Дж Мол Биол. 319 (2): 289–304. Дои:10.1016 / S0022-2836 (02) 00298-X. PMID 12051907. (Применение фага Т4 в биотехнологии для изучения взаимодействия белков)
- Tétart F .; Desplats C .; Кутателадзе М .; Monod C .; Ackermann H.-W .; Krisch H.M. (2001). «Филогения основных генов головы и хвоста обширных бактериофагов Т4-типа». J. Bacteriol. 183 (1): 358–366. Дои:10.1128 / JB.183.1.358-366.2001. ЧВК 94885. PMID 11114936. (Индикация распространенности последовательностей Т4-типа в дикой природе)
- Абедон С.Т. (2000). "Неясное происхождение Белоснежки и ее карликов-даже". Генетика. 155 (2): 481–6. ЧВК 1461100. PMID 10835374. (Историческое описание выделения Т4-подобных фагов Т2, Т4 и Т6)
- Акерманн Х.В., Криш Х.М. (1997). «Каталог бактериофагов Т4-типа». Arch. Вирол. 142 (12): 2329–45. Дои:10.1007 / s007050050246. PMID 9672598. Архивировано из оригинал 1 ноября 2001 г. (Почти полный список известных на тот момент Т4-подобных фагов)
- Monod C, Repoila F, Kutateladze M, Tétart F, Krisch HM (март 1997 г.). «Геном псевдо Т-четных бактериофагов, разнообразная группа, напоминающая Т4». J. Mol. Биол. 267 (2): 237–49. Дои:10.1006 / jmbi.1996.0867. PMID 9096222. (Обзор различных Т4-подобных фагов с точки зрения их геномов)
- Kutter E .; Гачечиладзе К .; Поглазов А .; Марусич Э .; Шнейдер М .; Aronsson P .; Напули А .; Портер Д .; Месянжинов В. (1995). «Эволюция T4-родственных фагов». Гены вирусов. 11 (2–3): 285–297. Дои:10.1007 / BF01728666. PMID 8828153. (Сравнение геномов различных Т4-подобных фагов)
- Карам, Дж. Д. и другие. 1994. Молекулярная биология бактериофага Т4. ASM Press, Вашингтон, округ Колумбия. (Вторая библия T4, перейдите сюда, а также Mosig and Eiserling, 2006, чтобы начать изучать биологию фага T4) ISBN 1-55581-064-0
- Эдди, С. Р. 1992. Интроны в T-четных бактериофагах. Кандидатская диссертация. Университет Колорадо в Боулдере. (Глава 3 содержит обзор различных T4-подобных фагов, а также выделение новых на тот момент T4-подобных фагов)
- Surdis, T.J. et al., Методы прикрепления бактериофагов, специфичные для T4, анализ, обзор.
- Мэтьюз, К. К., Э. М. Куттер, Г. Мозиг и П. Б. Бергет. 1983. Бактериофаг Т4. Американское общество микробиологии, Вашингтон, округ Колумбия. (Первая Библия Т4; не вся информация здесь дублируется в Караме. и другие., 1994; особенно см. вводную главу Doermann для исторического обзора T4-подобных фагов) ISBN 0-914826-56-5
- Рассел Р. Л. 1967. Видообразование среди T-четных бактериофагов. Кандидатская диссертация. Калифорнийский технологический институт. (Выделение ряда Т4-подобных фагов RB)
- Малыс Н, Нивинскас Р (2009). «Неканоническое расположение РНК в четных Т4 фагах: вмещает сайт связывания рибосомы в межцистронном соединении гена 26-25». Мол Микробиол. 73 (6): 1115–1127. Дои:10.1111 / j.1365-2958.2009.06840.x. PMID 19708923. (редкий тип регуляции трансляции, характерный для Т4)
- Kay D .; Файлдс П. (1962). «Гидроксиметилцитозин-содержащие и триптофан-зависимые бактериофаги, выделенные из городских сточных вод». J. Gen. Microbiol. 27: 143–6. Дои:10.1099/00221287-27-1-143. PMID 14454648. (Выделение Т4-подобного фага, включая фаг Ox2)