Eichstaettisaurus - Eichstaettisaurus
| Eichstaettisaurus | |
|---|---|
 | |
| Тип и единственный экземпляр Э. шредери | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Рептилии |
| Заказ: | Squamata |
| Clade: | Gekkonomorpha |
| Род: | †Eichstaettisaurus Кун, 1958 г. |
| Типовой вид | |
| Eichstaettisaurus schroederi (Бройли, 1938) | |
| Другие виды | |
| |
| Синонимы | |
Э. шредери
| |
Eichstaettisaurus (что означает «ящерица Eichstätt») является род из ящерицы от Позднеюрский период и Раннемеловой период Германии, Испании и Италии. С приплюснутой головой, ориентированными вперед и частично симметричными ступнями и высокими когтями, Eichstaettisaurus претерпел множество приспособлений к альпинистскому образу жизни, приближающемуся к гекконы. В типовой вид, Э. шредери, является одним из старейших и наиболее полных членов Squamata, известный по одному экземпляру, происходящему из Титонский в возрасте Зольнхофенский известняк Германии. Второй вид, Э. гулди, был описан с другого скелета, найденного в Матезе Горы Италии. Несмотря на то, что он очень похож на Э. шредери, он жил намного позже, во времена Альбианский сцена. Ископаемые остатки обоих видов демонстрируют исключительную сохранность благодаря отложению в с низким содержанием кислорода морская среда.
Первоначально назван как вид рода Ардеозавр к Фердинанд Бройли в 1938 г., Э. шредери имеет запутанную таксономическую историю. Первоначально его признали родственником ночные ящерицы пока исследования в 1960-х годах не выявили его и Ардеозавр как гекконы. Тем не мение, филогенетические анализы проведенные в 1990-х годах, предположили, что он был ближе к предку всех чешуйчатые. Различные противоречивые позиции были найдены до появления анализов, включающих больше видов и более точные данные, которые разрешили Э. шредери как близкий родственник гекконов в Gekkonomorpha. Однако позиция Ардеозавр в связи с Э. шредери остается неопределенным: один вид, A. digitatellus, был обнаружен как его близкий родственник в некоторых исследованиях, но другие виды A. brevipes было обнаружено, что они связаны более отдаленно.
Открытие и наименование
Э. шредери

В 1938 г. Фердинанд Бройли описал прекрасно сохранившийся образец ящерицы, сохранившийся вверх дном, из Юрский залежи горных пород в муниципалитете Винтерсхоф, Eichstätt, Германия. Образец попал во владение Баварской государственной коллекции палеонтологии и исторической геологии (BSPG), где его подготовил Л. Хаймерл. Он состоит из почти полного скелета, хотя и с раздробленным черепом, хвостом без хвоста после пятого позвонка (вероятно, в результате ампутация и замена на хрящ, а несколько позвонков сохранились только в виде отпечатков (но их наличие все еще можно было проверить по связанным ребрам). А контрплит к образцу тоже существует.[1] Сегодня известно, что месторождения горных пород относятся к Формация Альтмюльталь, а образец и его контрпланка хранятся под номерами доступа BSPG 1937 I 1a и 1b.[2]
Бройли отметил, что, хотя он не смог проверить это из первых рук. Кристиан Эрих Герман фон Мейер описание типовой образец из Ardeosaurus brevipes что он имел сходство с этим экземпляром, но также имел важные различия в морде и лобная кость форма, а также количество позвонков. Однако он не решился создать новый род из-за сохранности черепа. Таким образом, он отнес его к роду Ардеозавр с оговорками как новый вид, А.? Schröderi, названный в честь его коллеги Я. Шредера.[1] (Электрический ток конкретное имя является Schroederi: статьей 32.5.2.1 Международный кодекс зоологической номенклатуры, умляуты на немецком языке имена, опубликованные до 1985 года, должны быть удалены и заменены на букву "e" в качестве следующей буквы.)[3]
Роберт Хоффштеттер официально признал различие между А.? Schroederi и A. brevipes в 1953 г. и основал новый родовое имя, Broilisaurus. Тем не мение, Оскар Кун уже назвал стегоцефальный одноименная амфибия 1938 г .; таким образом, в 1958 году он установил еще одно родовое название вида, Eichstättisaurus. (Опять же, из-за ICZN текущее родовое имя Eichstaettisaurus.) В 1963 году Маргерит Кодуд-Мишель высказала мнение, что Э. шредери действительно представлял тот же вид, что и Homoeosaurus digitatellus, который ранее был назван нетиповым видом Гомеозавр в 1914 году Н.М.Гриером. Таким образом, она создала новую комбинацию Eichstaettisaurus digitatellus.[4][5][6]
В отличие от Кодуда-Мишеля, Хоффштеттер определил дигителлус как член Ардеозавр отдельно от Eichstaettisaurus в 1964 году. Хотя Кокуд-Мишель знала о статье Хоффштеттера, в 1965 году она повторила свой аргумент и не сделала попытки обратиться к Хоффштеттеру. Хоффштеттер ответил в 1966 году, назвав синонимию Кокуд-Мишель «невыносимой». Он отметил, что это повлечет за собой пересмотр диагностические характеристики из Eichstatettisaurus до такой степени, что он будет практически неотличим от Ардеозавр, что противоречило бы ей семейный уровень разделение двух.[6] Последующая литература сохранила Eichstaettisaurus schroederi и Ardeosaurus digitatellus как отдельные, хотя им уделялось мало внимания, пока Тиаго Симойнс и его коллеги не опубликовали переописание обоих в 2017 году.[2]
Э. гулди

На протяжении многих лет, Eichstaettisaurus был известен по единственному экземпляру Э. шредери до того как Меловой Престарелые останки были найдены в других частях Европы. В 2000 г. Сьюзан Эванс и его коллеги описали небольшой образец ящерицы в Берриасский –Валанжинский старый район Монсек в Каталония, Испания. Несмотря на плохую сохранность, этот экземпляр явно отличался от более распространенных здесь ящериц. Мейазавр; Эванс и его коллеги предположили, что он имеет сходство с Eichstaettisaurus.[7]
В 2004 году Эванс и его коллеги сообщили о еще более молодых останках Eichstaettisaurus, который возник из Альбианский в возрасте Pietraroja Plattenkalk в районе Пьетрароя, который находится в Матезе Горы южной Италии. Экземпляр, внесенный в каталог как MPN 19457, сформировал типовой образец нового вида, Э. гулди, который они назвали в честь покойного биолога Стивен Джей Гулд; Это скелет, сохранившийся снизу вверх, который включает череп, части позвоночника и конечности. Второй образец состоит из разрозненного черепа, который сохранился как часть содержимого кишечника неопределенного ринхоцефальный каталогизирован как MPN A01 / 82. Оба экземпляра хранятся в Музее палеонтологии на Неаполитанский университет (MPN).[4]
Описание
Eichstaettisaurus был относительно небольшим животным. Э. шредери имел длину от носа до отверстия (измерялась от кончика морды до отверстия клоака ) 94 мм (3,7 дюйма),[2] пока Э. гулди имел длину носа-отверстия 56 мм (2,2 дюйма).[4]
Череп
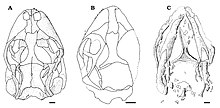
Eichstaettisaurus можно определить по уплощенному черепу с короткой, тупой и округлой мордой. Э. шредери были большие ноздри. Зубные кости рыла, предчелюстная кость и верхняя челюсть, вероятно, были связаны группой мягких тканей в Э. шредери. На предчелюстной кости было 6 или 7 зубов в Э. гулди, но число в Э. шредери неизвестно; на верхней челюсти было примерно 22 зуба в Э. шредери, и не менее 30 в Э. гулди. Зубы у обоих видов были мелкие, узкие, конические, одно-заостренный; у них был плевродонт имплантация, означающая, что они были сращены с внутренней стороной челюстной кости. Нравиться Ардеозавр, оба вида не имели слезная кость, обычно часть глазницы. Также в глазнице, лобной и заглазничные кости были отдельными в отличие от Ардеозавр, а задний проекция из последних костей были относительно широкими. В Э. шредери, контакт между этими костями был дальше, чем у большинства ящериц. В чешуйчатые кости были тонкими и спускались вниз, чтобы соответствовать квадратные кости, как и у большинства ящериц; в отличие от Игуания и Teiidae однако прогнозов на повышение у них не было. У живых гекконов отсутствуют заглазничные и чешуйчатые кости, а также они относительно короче. скуловые кости чем Eichstaettisaurus. На нёбо из Э. шредери, эктоптеригоидные кости перекрывали крыловидные кости в их тылу.[2][4]
В отличие от A. brevipes, Э. шредери отсутствовал костный орнамент и остеодермы на крыша черепа. У обоих видов лобные кости срослись и стали более узкими между глазницами, чем спереди. Нисходящие выступы на лобных костей, известные как субольфакторные отростки, были хорошо развиты у обоих видов, но встречались на средней линии только у Э. шредери. В шов между лобной и теменные кости был выпуклым в Э. шредери и вогнуться Э. гулди, В отличие от Ардеозавр где было прям. Э. гулди и у большинства других ящериц теменные кости срослись, но они были спарены и не срослись в Э. шредери. Теменные кости обоих видов без задних выступов и затылочной кости. ямки. В Э. шредеривнешние края теменных костей загнуты внутрь, а задние выступы, известные как надвисочные отростки, были короткими, широко разделенными и имели углубления. Также в Э. шредери, пара гербов присутствовала на надзатылочная кость черепной коробки, которые, вероятно, были отпечатаны полукружные каналы из-за уменьшенного черепа окостенение.[2][4]
Позвонки и ребра
По сравнению с большинством игуанцев, геккотан, и сцинкоморфан ящерицы, у которых обычно 24-26 позвонки напротив крестец (или бедро), Э. шредери было 31, в том числе 7 шейные позвонки и примерно 24 задние позвонки.[2] Э. гулди имело 6–7 шейных позвонков и неизвестное количество задних позвонков.[4] В Э. гулди, второй (ось ) и третий шейный позвонок были соединены между собой элементами, известными как intercentra.[4] В Э. шредери, большая часть шейных позвонков отсутствовала шейные ребра, за исключением двух последних. Восходящий нервные отростки на задних позвонках были маленькие, а последние несколько задних позвонков отсутствовали ребра в духе поясничных позвонков. Зигосфен-зигантрум сочленения присутствуют на задних позвонках обоих видов. Ребра были одноголовыми и имели круглые шарнирные поверхности.[2] В Э. гулди, то центр (основные тела) хвостовых позвонков широкие, цилиндрические, амфицеллярный (вогнутые с обоих концов), а отверстие сильно развито поперечные отростки.[4]
Конечности и пояса конечностей

Eichstaettisaurus имел хорошо развитый акромион процесс, или приложение для ключица, на лопатка (лопатка). Ключицы Э. шредери не наклонился вперед, в отличие от большинства членов Autarchoglossa, а также отсутствовала задняя проекция. Как в Ардеозавр, оба вида Eichstaettisaurus имел относительно крепкие передние конечности.[2][4] В Э. шредери, то олекранон процесс локтевая кость был хорошо развит.[2] В Э. гулди, то кости запястья вероятно, не полностью окостенели.[4] Пятизначная рука Э. шредери отверстие 2, 3, 4, 5 и 3 фаланги (кости пальцев) на каждом пальце; предпоследние фаланги от кончиков были относительно длинными.[2] Э. гулди на четвертом пальце имел 4 фаланги, когти короткие, глубокие и острые, с хорошо развитым сгибателем. бугорки.[4]
в таз из Э. шредери, расширение лобковые волосы в сросшийся лобковый фартук был относительно узким; лобковые волосы также не имели передних выступов, известных как лобковые бугорки. В искья были увеличены и сильно контактировали друг с другом, а заднее расширение лезвия подвздошная кость было долго. Обе Eichstaettisaurus виды имели крепкие задние конечности, по длине близкие к передним. В бедренная кость имел прямой стержень, в отличие от большинства других ящериц, где он сигмовидный. В ногу первая плюсневая кость был немного короче второй-четвертой плюсневых костей. Необычно пятая плюсневая кость была намного короче и крючковатой. В Э. шредеринога была пятипалой, как и руки, с 2, 3, 4, 5 и 4 фалангами на каждом пальце, и предпоследние фаланги и когти были похожи. Э. шредери разделял две особенности когтей на ногах с живыми Токайский геккон: когти с расширенными нижними концами; а сочленения между фалангами были простыми вогнуто-выпуклыми суставами, а не двояковидный (т.е. основанный на двух закругленных выступах на концах фаланг), в отличие от большинства других ящериц.[2][4]
Классификация

В филогенетический история Eichstaettisaurus был бурным, как у Ардеозавр, что было следствием как неполных данных, так и их примитивной формы.[2][8] фон Мейер не предоставил размещение для Ардеозавр когда он назвал род; Ричард Лидеккер и Карл Альфред фон Циттель поместил это и Гомеозавр в Rhynchocephalia, Барон Франц Нопца размещен Ардеозавр как близкий родственник сцинки, и Чарльз Кэмп поместил его рядом с Gekkonidae.[9][10][11] Судя по хорошо развитым краям глазниц и надвисочных окон на черепе у типового экземпляра Э. шредери, Бройли согласился с выводом Нопса в 1938 году. В пределах Scincomorpha он отметил, что амфикоелозные позвонки Ардеозавр (включая Э. шредери) лучше всего напоминал Xantusiidae (ночные ящерицы), как и у большинства других сцинков, имеют прцелевидные (выпуклые на задней поверхности) позвонки.[1]
При разделении Schroederi в род Broilisaurus, Хоффштеттер сохранил его в составе Xantusiidae в 1953 году, но заметил, что его череп удивительно напоминает череп геккона. Поскольку ксантусииды и гекконы были помещены в две основные признанные ветви ящериц, Autarchoglossa и Аскалабота Хоффштеттер предположил, что он был близок к предкам ящериц.[5] После переименования его в EichstaettisaurusКун основал семейство Eichstaettisauridae, все еще относящееся к Xantusiidae. В 1961 году, а затем в 1963 году Кодуд-Мишель согласился с Кэмпом в том, что Ardeosauridae и Eichstaettisauridae на самом деле были гекконами, а не ксантусиидами, что объясняется значительными различиями в посткраниальном скелете, и оба помещались в Gekkonomorpha.[5] Хоффштеттер принял это определение в 1966 году,[6] как и Ричард Эстес в 1983 году.[2]
Тем не мение, филогенетические анализы под руководством Виктора Рейносо в 1996 г. (не опубликовано), Эванса и Луиса Хавьера Барбадилло в 1998 и 1999 гг., а также Эванса и Даниэля Чуре в 1998 г. Eichstaettisaurus и Ардеозавр были намного больше базальный (рано расходящиеся) чешуйчатые. Вместе с Баваризавр, Scandensia, и Hoyalacerta, эти двое выпали за пределы группа короны Squamata. Это сделало бы их стеблевыми-чешуйчатыми, то есть не потомками общего предка всех ныне живущих чешуйчатых.[2][12][13] Однако в анализе 1999 г. Эванс и Барбадилло признали, что позиция Eichstaettisaurus был нестабильным из-за отсутствия данных (только 44% характеристик, используемых в их анализе, могли быть закодированы для него), и одна из их филогенетические деревья поместил его как сестринская группа к Склероглосса среди чешуек кроны.[14] Эванс и его коллеги обнаружили Э. гулди быть сестринской группой Э. шредери в 2004 году и восстановили одинаковое положение для обоих.[4]
Последующий филогенетический анализ Эванса и его коллег выявил множество противоречивых позиций по Eichstaettisaurus и Ардеозавр. В 2005 году Эванс и Юань Ван провели два анализа, один на основе анализа Майкла Ли 1998 года, а другой - на основе анализа Эванса и Чуре 1998 года, но они добавили: Далингозавр, Дорсетизавр, и Парвираптор как для. Один найден Eichstaettisaurus как сестринская группа группы, содержащей Xantusiidae, Gekkonidae, Pygopodidae, и Амфисбания, с Ардеозавр быть близко к Lacertidae и Teiioidea; другой объединился Eichstaettisaurus с Далингозавр и Xenosauridae в Anguimorpha, с Ардеозавр как сестринская группа Игуании.[15] После того, как Эванс, Ван и Чун Ли добавили Ябейнозавр к тем же анализам, было найдено Eichstaettisaurus, Ардеозавр, Ябейнозавр, и Парвираптор сформировать сестринскую группу Склероглоссу, в то время как другая выздоровела Ардеозавр как стебель-чешуйчатый и Eichstaettisaurus, Hoyalacerta, Парвираптор, и Scandensia сформировать группу среди склероглоссов.[16]
Филогенетический анализ 2006 г., проведенный Джеком Конрадом и Марк Норелл аналогично найдено Eichstaettisaurus как можно ближе к склероглоссу.[17] В 2008 году Конрад опубликовал большой филогенетический анализ чешуек, включающий 222 таксоны и 363 характеристики, в которых была предпринята попытка решить проблему неполного отбора таксонов (т.е. недостаточное количество включенных таксонов), вызывающих филогенетическую нестабильность. Включать Eichstaettisaurus, он объединил данные из Э. шредери и Э. гулди. Он обнаружил, что оба Eichstaettisaurus и Ардеозавр, вместе с Баваризавр, входили в основную группу Scleroglossa. Наряду с коронной группой Scleroglossa они включали Сцинкогеккономорфа.[18] Арнау Болет и Эванс провели два анализа, основанные на анализе Конрада, для описаний 2010 и 2012 гг. Педреразавр и Джукарасепс, и восстановил аналогичные позиции для Eichstaettisaurus. Однако первый содержал политомия так как отношения между Игуанией, Педреразавр, группа, содержащая Ардеозавр, Ябейнозавр, и Сакурасавр, и все другие чешуйки не могут быть решены.[19][20]
Жак Готье и его коллеги опубликовали еще один большой филогенетический анализ в 2012 году, содержащий 192 таксона и 976 характеристик. Как и Конрад, они также объединили данные из Э. шредери и Э. гулди. Они восстановили более производительную позицию для Eichstaettisaurus в составе стволовой группы Gekkota вместе с безымянным экземпляром AMNH FR 21444; в частности, они отметили, что пропорции конечностей сильно напоминают геккотаны.[8] После добавления Ardeosaurus digitatellus и перекодирование Eichstaettisaurus включать только данные из Э. шредери, Симойнс и его коллеги нашли Э. шредери в аналогичном положении, хотя и в политомии с A. digitatellus и AMNH FR 21444. Они идентифицировали ряд признаков геккотана: парные теменные кости; закрытый Меккелевская бороздка на нижней челюсти; маленькие конические зубы верхней челюсти; короткая тупая морда и приплюснутый череп; эктоптеригоиды перекрывают крыловидные кости; два крыловидных тела и небные кости быть широко разделенными; широкий шов отделение крыловидных отростков и небных костей друг от друга; и поясничные позвонки.[2]
В 2018 году Матеуш Таланда добавил Ardeosaurus brevipes к анализу Готье и его коллег (но не включает исправления Симойнса и его коллег). Он нашел Eichstaettisaurus в том же месте рядом с Нореллиус, но и выздоровел A. brevipes как сцинкоморф, тесно связанный с сцинками (в отличие от стебл-геккотанской позиции A. digitatellus). Несмотря на различное филогенетическое положение, Таланда заметил, что эти два вида различаются только шириной их теменной кости. Он предположил, что неполные данные из-за плохой сохранности A. digitatellus могли повлиять на эти результаты, и поэтому он сохранил его в роде Ардеозавр.[21] Другой анализ, проведенный Симойнсом, Таландой и коллегами в 2018 году, на этот раз с использованием уточненных данных Симойнса и его коллег, обнаружил аналогичную позицию для Э. шредери но более базальный для A. brevipes в Autarchoglossa.[22] В 2019 году Владимир Алифанов рассмотрел оба Eichstaettisaurus и Ардеозавр как сцинкоморфы, тесно связанные с Карусия но не к Xenosauridae, но филогенетический анализ не проводился.[23]
Ниже, филогенетические деревья из двух основных филогенетических анализов, включая Eichstaettisaurus показаны: один основан на исследовании Конрада (Bolet & Evans, 2012, с отношениями внутри Scleroglossa из Conrad, 2008),[18][20] и один основан на исследовании Готье и его коллег (Simões et al., 2018).[22]
Топология A: Bolet & Evans (2012) со склероглоссой от Конрада (2008)[18][20]
| Squamata |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Топология B: Simões и другие. (2018)[22]
| Squamata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология

Современные гекконы необычны среди ящериц тем, что пальцы их конечностей относительно симметричны по длине и расставлены широкой дугой; Напротив, у других ящериц пальцы обычно почти параллельны друг другу, особенно на лапах. Узор, наблюдаемый у гекконов, облегчает захват, когда тело находится в различной ориентации, поскольку он распределяет клей щетинки (щетина) на подушечках пальцев ног, позволяя первым и последним пальцам располагаться напротив друг друга.[24][25] Eichstaettisaurus schroederi имел две характеристики, которые способствовали симметрии стопы: его первая плюсневая кость была длинной относительно ее третьей плюсневой кости (третья плюсневая кость была всего в 1,41 раза длиннее первой, в отличие от 2-х раз у типичной ящерицы) и четвертая плюсневая кость была относительно короткий. В 2017 году Симойнс и его коллеги заметили, что Э. шредери имел более сильную симметрию стопы, чем Ardeosaurus digitatellus, и они пришли к выводу, что ноги обоих, вероятно, были направлены вперед дальше, чем у других ящериц. Это также подтверждается отсутствием двояковыпуклых сочленений в фалангах, которые являются адаптацией для обращенных наружу стоп, которые сопротивляются смещению в горизонтальной плоскости, обеспечивая сгибание в вертикальной плоскости.[2]
Симойнс и его коллеги также определили несколько других характеристик Э. шредери, которые предполагают, что сканирующий (основанный на лазании) образ жизни современных гекконов возник раньше, чем считалось ранее. Его высокие когти и удлиненные фаланги на кончиках пальцев на пальцах - черты, которые сильно коррелировали со сканирующим образом жизни ящериц.[26][27][28][29][30] В частности, высокие когти могли обеспечивать более длинные рычаги для сухожилий сгибателей, которые втягивают их, таким образом улучшая их силу захвата.[31] Между тем, его относительно короткие конечности и сплющенное тело могли улучшить характеристики лазания за счет понижения его центра тяжести, как это предполагалось для токайского геккона.[31] но эта функция может не соотноситься со сканирующим образом жизни.[32] Наконец, его конечности такой же длины могли иметь улучшенный захват, как в остроногая каменная ящерица; Значение этого признака может быть уменьшено из-за липких подушечек ног гекконов и отсутствия корреляции между соотношением длины конечностей и сканированием Lacertidae.[2][32]
В 2004 году Эванс и его коллеги также обсудили образ жизни Э. гулди в свете его телесных пропорций.Они указали на предыдущее исследование того, как различия в пропорциях среди видов снежных сцинков (Каринаскинкус ) коррелировали с использованием среды обитания: короткие туловища и длинные задние конечности коррелировали с лазанием по скалам (у крупных видов) и лазанием по деревьям (у мелких видов), тогда как длинные туловища и короткие задние конечности коррелировали с обитанием на земле.[33] Эванс и его коллеги обнаружили, что Э. гулди По своим пропорциям был наиболее близок к наземным видам, которые имеют низкую скорость бега и относительно плохо лазают. Тем не менее они признали, что плоские тела и высокие когти Э. гулди были приспособлены для цепляния за грубые поверхности. Они предложили гибридный образ жизни для Э. гулди: медлительная наземная ящерица, способная лазить по скалам и прятаться в расщелинах от хищников, таких как ринхоцефалы.[4]
Палеоэкология
Э. шредери

Каменные отряды в Винтерсхофе, которые произвели единственный известный образец Э. шредери являются частью Солнхофенские известняки южной Германии, которые хорошо известны своими исключительно сохранившимися окаменелостями.[2] Исторически сложилось так, что стратиграфия известняков Зольнхофена были предметом значительной путаницы, поскольку разнообразие сред и структур осадконакопления способствовало неразберихе.[34] Теперь считается, что они состоят из нескольких геологические образования в Группа Weißjura: в самом строгом смысле они состоят из Формация Альтмюльталь к северо-западу от Ингольштадт (включая населенные пункты вокруг Айхштетта) и Пейнтен Формирование к северо-востоку от Ингольштадта. В пределах формации Альтмюльталь карьер Винтерсхоф является частью верхней пачки Айхштетт, которая с точки зрения аммонит биостратиграфия лежит в Euvirgalithacoceras eigeltingense -β горизонт между Lithacoceras riedense и Hybonoticeras hybonotum подзоны.[35][36] Отложения формации Альтмюльталь, датируемые нижним Титонский сцена из Юрский период, состоит из Plattenkalk (очень мелкозернистый известняк грязи на основе, такие как микрит ), образуя ровные тонкие слои толщиной около 1 сантиметра (0,39 дюйма), которые обычно не имеют доказательств биотурбация (нарушение живыми организмами).[37][38][39]
В течение титона платтенкалк формации Альтмюльталь откладывался в океанические бассейны (так называемый "ваннен") в теплом мелком море, окружающем архипелаг.[39] Эти бассейны, которые, возможно, были лагуны, имели палеошироту примерно 34 ° с.ш. и располагались на северной окраине Тетис Океан. Окаменелости донных животных, таких как хрупкие звезды и брюхоногие моллюски практически отсутствуют, что свидетельствует о неблагоприятных для жизни условиях на морском дне; это могло быть вызвано одним из нескольких факторов, включая гиперсоленость, кислородное истощение, или накопление токсичных сероводород.[38][40] Эти условия были ответственны за исключительную сохранность окаменелостей, подобных ископаемым. Э. шредери,[41] которые преимущественно переносились с окружающей земли во время сильных дождей и штормов.[42] На суше отсутствие папоротники и преобладание сухоадаптированных хвойные породы предполагаю, что климат был полузасушливый.[43]

Обе Ardeosaurus brevipes и A. digitatellus были обнаружены в Wintershof с Э. шредери,[2] вместе с Homoeosaurus maximiliani и ринхоцефал Плеврозавр голдфусси.[5] Также из Wintershof известны птерозавр Rhamphorhynchus muensteri,[44] то крокодиловидный Alligatorellus bavaricus,[45] и различные водные животные: рыбы Anaethalion angustus,[46] Ascalabos voithii,[47] Aspidorhynchus acutirostris,[48] Belonostomus spyraenoides,[49] Caturus giganteus,[50] Gyrodus круговой,[51] Macrosemius rostratus, Palaeomacrosemius thiollieri,[52] Propterus elongatus,[53] и Zandtfuro tischlingeri;[54] то ангельская акула Pseudorhina alifera;[55] кальмар колеоиды Acanthoteuthis problematica,[56] Belemnotheutis mayri,[57] и Plesioteuthis prisca;[58] то криноид Saccocoma tenella, что очень часто встречается в отложениях Солнхофена;[59] креветка Дуса Реши;[60] и подковообразный краб Mesolimulus walchi.[61] Соседние карьеры произвели Образец Айхштетта из Avialan динозавр Литографический археоптерикс,[62] и птерозавры Aerodactylus scolopaciceps, Germanodactylus cristatus, и возможно Cycnorhamphus.[63]
Э. гулди

Местность Пьетрароя, откуда Э. гулди экз., образован обнажением платтенкалка в верхней части скальный склон («Чивита ди Пьетрароя»), расположенный на восточной окраине гор Матезе. Эти горы не начинали формироваться до Миоцен, с большинством тектоническое поднятие который привел их к их нынешнему положению, произошел между Плиоцен и Плейстоцен. На местности есть две отчетливые пластинки; нижний слой более крупнозернистый и в основном лишен окаменелостей, а верхний слой более илистый и является источником большинства окаменелостей. Две пласты разделены лагунными известняками. Платтенкалк образован слоями Packstone и Грейнстоун толщиной 2–25 см (0,79–9,84 дюйма), которые чередуются со слоями аргиллит, чокнутый камень, и мергель; верхняя кровать также отличается наличием спикулы губки, черт, битум, и копролиты.[4][64]
На основе фораминиферы, местонахождение Пьетароя датируется Альбианский эпоха раннего мела. За это время платтенкалк Пьетрарои был бы заложен на мелководье. карбонатная платформа недалеко от небольшого острова. Первоначально предполагалось, что осадочная среда была лагуной,[65] но он был интерпретирован как восточный подводный канал он постепенно заполнялся в течение апта, основываясь на закономерностях в расположении окаменелостей, водных течениях и переносе отложений. Отсутствие биотурбации также указывает на бескислородную среду, возможно связанную с глобальным океаническим аноксическое событие. Некоторые окаменелости хорошо сохранились, в то время как другие были почти уничтожены, что позволяет предположить, что они произошли из разных мест, с такими наземными животными, как Э. гулди заносится в канал потоками пресной воды.[4][64] Климат был бы тропический к субтропический.[65]

В добавление к Э. гулди и ринхоцефал MPN A01 / 82, др. лепидозавры из Пьетрарои включают ящериц Chometokadmon fitzingeri и Костазавр рускони, а ринхоцефальный Derasmosaurus pietraroiae а также третий безымянный молодой особь.[4][66][67] Наиболее известная окаменелость из Пьетрарои - окаменелость Compsognathid динозавр Сципионикс самнитический, который сохраняет мягкие ткани, в том числе внутренние органы.[68] Среди других рептилий есть два крокодиловидных образца, принадлежащих к Pietraroiasuchus ormezzanoi,[69] и альбанерпетонтид амфибия Celtedens megacephalus.[70] Многие рыбы известны из Пьетрарои, наиболее распространенной из которых является пикнодонтид ранее назывался Coelodus costae но теперь известен как Gregoriopycnodus bassanii.[71] Другие включают Анаэталион робустус, Belonostomus crassirostris, Caeus leopoldi, Cavinichthys pietrarojae, Хироцентриты coroninii, родственники Diplomystus brevissimus и Elopopsis fenzii, Гемиелопоз гиббус, Ionoscopus petrarojae, Italophiopsis derasmoi, разновидность Лепидозы, Notagogus pentlandi, Pleuropholis decastroi, Propterus scacchii, и Sauropsidium laevissimum.[72][73][74][75][76] Хрящевые рыбы включали акулу-ангела Phorcynis и луч Ринобатус обтусатус.[72] Включены беспозвоночные рудисты, двустворчатые моллюски, морские улитки рода Неринея, креветки и морские звезды.[64][65]
Рекомендации
- ^ а б c Бройли, Ф. (1938). "Ein neuer fund von?Ардеозавр Х. фон Мейер " [Новая находка?Ардеозавр Х. фон Мейер] (PDF). Sitzungsberichte der Mathematisch-Naturwissenschaftlichen Abteilung der Bayerischen Akademie der Wissenschaften zu München: 97–114.
- ^ а б c d е ж грамм час я j k л м п о п q р s Simões, T.R .; Caldwell, M.W .; Nydam, R.L .; Хименес-Уидобро, П. (2017). «Остеология, филогения и функциональная морфология двух видов юрских ящериц и ранняя эволюция скансориальности у гекконов». Зоологический журнал Линнеевского общества. 180 (1): 216–241. Дои:10.1111 / zoj.12487.
- ^ Snitting, D .; Блом, Х. (2009). «Исправление названий таксонов, содержащих диакритические знаки - примеры палеозойских позвоночных». Журнал палеонтологии позвоночных. 29 (1): 269–270. Дои:10.1080/02724634.2009.10010377.
- ^ а б c d е ж грамм час я j k л м п о п q Evans, S.E .; Raia, P .; Барбера, К. (2004). «Новые ящерицы и ринхоцефалы из нижнего мела южной Италии» (PDF). Acta Palaeontologica Polonica. 49 (3): 393–408.
- ^ а б c d Кодуд-Мишель, М. (1963). "Les rhynchocéphales et les sauriens des calcaires lithographiques (Jurassique supérieur) Западной Европы" [Ринхоцефалы и ящеры из литографических известняков (верхняя юра) Западной Европы]. Новые архивы Лионского музея естественной истории. 7: 143–185.
- ^ а б c Хоффштеттер Р. (1966). "Предложение жанров Ардеозавр et Eichstaettisaurus (Reptilia, Sauria, Gekkonoidea) du Jurassique Supèrieur de Franconie " [О родах Ардеозавр и Eichstaettisaurus (Reptilia, Sauria, Gekkonoidea) из верхней юры Франции]. Bulletin de la Société Géologique de France. 8 (4): 592–595. Дои:10.2113 / gssgfbull.S7-VIII.4.592.
- ^ Evans, S.E .; Ruiz, A.L .; Рей, Дж. (2000). «Ящерица из раннего мела (берриас-валанжин) в Монсеке, Каталония, Испания». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 215: 1–15. Дои:10.1127 / njgpa / 215/2000/1.
- ^ а б Gauthier, J.A .; Kearney, M .; Maisano, J.A .; Rieppel, O .; Behlke, A.D.B. (2012). «Сборка плоского древа жизни: перспективы фенотипа и летописи окаменелостей». Бюллетень музея естественной истории Пибоди. 53 (1): 3–308. Дои:10.3374/014.053.0101.
- ^ Лидеккер, Р. (1888). «Отряд ринхоцефалии». Каталог ископаемых рептилий и амфибий в Британском музее (естественная история). 1. Лондон: Британский музей естественной истории. С. 290–301. Дои:10.5962 / bhl.title.61848.
- ^ Нопса, Ф. (1908). "Zur Kenntnis der fossilen Eidechsen" [К сведениям об ископаемых ящерицах]. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients. 21: 33–62.
- ^ Кэмп, К. (1923). «Классификация ящериц». Бюллетень Американского музея естественной истории. 48: 289–481. HDL:2246/898.
- ^ Evans, S .; Барбадилло, Л. Дж. (1998). «Необычная ящерица (Reptilia: Squamata) из раннего мелового периода Лас-Ойяс, Испания». Зоологический журнал Линнеевского общества. 124 (3): 235–265. Дои:10.1006 / zjls.1997.0139.
- ^ Evans, S.E .; Чуре, округ Колумбия (1998). "Черепа парамацеллодидных ящериц из юрской формации Моррисон в Национальном памятнике динозавров, штат Юта". Журнал палеонтологии позвоночных. 18 (1): 99–114. Дои:10.1080/02724634.1998.10011037.
- ^ Evans, S.E .; Барбадилло, L.J. (1999). "Коротконогая ящерица из нижнего мела Испании". Специальные статьи по палеонтологии. 60: 73–85.
- ^ Evans, S.E .; Ван, Ю. (2005). "Раннемеловая ящерица Далингозавр из Китая" (PDF). Acta Palaeontologica Polonica. 50: 725–742.
- ^ Evans, S.E .; Wang, Y .; Ли, К. (2005). "Раннемеловой род ящериц Ябейнозавр из Китая: разгадывая загадку ». Журнал систематической палеонтологии. 3 (4): 319–335. Дои:10.1017 / с1477201905001641.
- ^ Conrad, J.L .; Норелл, М. (2007). «Полный позднемеловой игуан (Squamata: Reptilia) из Гоби и определение новой клады игуан». Американский музей Novitates. 3584: 1–47. Дои:10.1206 / 0003-0082 (2007) 3584 [1: ACLCIS] 2.0.CO; 2. HDL:2246/5877.
- ^ а б c Конрад, Дж. Л. (2008). «Филогения и систематика чешуек (Reptilia) на основе морфологии». Бюллетень Американского музея естественной истории. 310: 1–182. Дои:10.1206/310.1. HDL:2246/5915.
- ^ Болет, А .; Эванс, С. (2010). «Новая ящерица из раннего мела Каталонии (Испания) и мезозойские ящерицы Пиренейского полуострова». Меловые исследования. 31 (4): 447–457. Дои:10.1016 / j.cretres.2010.06.002.
- ^ а б c Болет, А .; Эванс, С. (2012). «Крошечная ящерица (Lepidosauria, Squamata) из нижнего мела Испании». Палеонтология. 55 (3): 491–500. Дои:10.1111 / j.1475-4983.2012.01145.x.
- ^ Таланда, М. (2018). «Исключительно сохранившийся юрский сцинк предполагает диверсификацию ящериц, предшествовавшую фрагментации Пангеи». Палеонтология. 61 (5): 659–677. Дои:10.1111 / pala.12358.
- ^ а б c Simões, T.R .; Caldwell, M.W .; Таланда, М .; Бернарди, М .; Palci, A .; Вернигора, О .; Бернардини, Ф .; Mancini, L .; Найдам, Р.Л. (2018). «Происхождение чешуек, обнаруженное ящерицей среднего триаса из итальянских Альп». Природа. 557 (7707): 706–709. Bibcode:2018Натура.557..706S. Дои:10.1038 / s41586-018-0093-3. PMID 29849156.
- ^ Алифанов, В. (2019). «Ящерицы семейств Eoxantidae, Ardeosauridae, Globauridae и Paramacellodidae (Scincomorpha) из апта – альба Монголии». Палеонтологический журнал. 53 (1): 74–88. Дои:10.1134 / s0031030119010039.
- ^ Рассел, А.П. (1986). «Морфологические основы весовой нагрузки на сканерах токайского геккона (Reptilia: Sauria)». Канадский журнал зоологии. 64 (4): 948–955. Дои:10.1139 / z86-144.
- ^ Russell, A.P .; Bauer, A.M .; Ларойя, Р. (1997). «Морфологические корреляты вторично-симметричных ног геккотанских ящериц». Журнал зоологии. 241 (4): 767–790. Дои:10.1111 / j.1469-7998.1997.tb05747.x.
- ^ Зани, П.А. (2000). «Сравнительная эволюция морфологии когтей и пальцев ящерицы и способности цепляться». Журнал эволюционной биологии. 13 (2): 316–325. Дои:10.1046 / j.1420-9101.2000.00166.x.
- ^ Tulli, M.J .; Cruz, F.B .; Herrel, A .; Vanhooydonck, B .; Абдала, В. (2009). «Взаимодействие между морфологией когтей и использованием микропредприятий у неотропических игуанских ящериц». Зоология. 112 (5): 379–392. Дои:10.1016 / j.zool.2009.02.001. PMID 19632100.
- ^ Crandell, K.E .; Herrel, A .; Sasa, M .; Losos, J.B .; Осень, К. (2014). "Палка или захват? Совместное развитие липких подушечек пальцев ног и когтей в Анолис ящерицы ". Зоология. 117 (6): 363–369. Дои:10.1016 / j.zool.2014.05.001. PMID 25069967.
- ^ Kavanagh, K.D .; Шовал, О .; Уинслоу, Б. Б.; Alon, U .; Лири, Б.П .; Кан, А .; Табин, С.Дж. (2013). «Предвзятость развития в эволюции фаланг». Труды Национальной академии наук. 110 (45): 18190–18195. Bibcode:2013PNAS..11018190K. Дои:10.1073 / pnas.1315213110. ЧВК 3831474. PMID 24151335.
- ^ Rothier, P.S .; Brandt, R .; Колсдорф, Т. (2017). «Экологические ассоциации аутоподиальной остеологии у неотропических гекконов». Журнал морфологии. 278 (3): 290–299. Дои:10.1002 / jmor.20635. PMID 28112828.
- ^ а б Рассел, А.П. (1975). "Вклад в функциональный анализ стопы токайцев, Гекко геккон (Рептилии: Gekkonidae) ". Журнал зоологии. 176 (4): 437–476. Дои:10.1111 / j.1469-7998.1975.tb03215.x.
- ^ а б Vanhooydonck, B .; Ван Дамм, Р. (1999). «Эволюционные взаимосвязи между формой тела и использованием среды обитания у ящериц-ящериц» (PDF). Исследования эволюционной экологии. 1: 785–805.
- ^ Melville, J .; Суэйн, Р. (2000). «Эволюционные взаимосвязи между морфологией, производительностью и открытостью среды обитания у ящериц рода. Нивеосцинкус (Scincidae: Lygosominae) ". Биологический журнал Линнеевского общества. 70 (4): 667–683. Дои:10.1111 / j.1095-8312.2000.tb00222.x.
- ^ Швайгерт, Г. (2007). «Биостратиграфия аммонитов как инструмент для датировки литографических известняков верхней юры Южной Германии - первые результаты и открытые вопросы». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 245 (1): 117–125. Дои:10.1127/0077-7749/2007/0245-0117.
- ^ Niebuhr, B .; Пюрнер, Т. (2014). "Plattenkalk und Frankendolomit - Lithostratigraphie der Weißjura-Gruppe der Frankenalb (außeralpiner Oberjura, Bayern)". Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften. 83: 5–72.
- ^ Rauhut, O.W.M .; Tischlinger, H .; Фот, К. (2019). "Неархеоптеригидный птичий теропод из поздней юры на юге Германии". eLife. 8: e43789. Дои:10.7554 / eLife.43789. ЧВК 6516837. PMID 31084702.
- ^ Mönnig, E .; Franz, M .; Швайгерт, Г. (2018). «Der Jura in der Stratigraphischen Tabelle von Deutschland (STD 2016)» [Стратиграфическая карта Германии (STD 2016): юрский период]. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften. 169 (2): 225–246. Дои:10.1127 / zdgg / 2018/0148.
- ^ а б Munnecke, A .; Westphal, H .; Кёльбл-Эберт, М. (2008). «Диагенез платтенкалка: примеры из района Зольнхофен (верхняя юра, юг Германии)». Седиментология. 55 (6): 1931–1946. Bibcode:2008Седим..55.1931M. Дои:10.1111 / j.1365-3091.2008.00975.x.
- ^ а б Уилкин, Дж. (2020). «Южногерманский Платтенкалкс». Геология сегодня. 36 (1): 27–32. Дои:10.1111 / gto.12288.
- ^ Виол, Г. (1994). «Рыбная тафономия платтенкалка Солнхофена - подход к реконструкции палеосреды». Geobios. 27: 81–90. Дои:10.1016 / с0016-6995 (94) 80023-5.
- ^ Seilacher, A .; Reif, W.E .; Вестфаль Ф. (1985). «Седиментологические, экологические и временные закономерности ископаемых лагерштеттенов». Философские труды Лондонского королевского общества B. Биологические науки. 311 (1148): 5–24. Bibcode:1985РСПТБ.311 .... 5С. Дои:10.1098 / rstb.1985.0134.
- ^ Beardmore, S.R .; Lawlor, E .; Hone, D.W.E. (2017). «Использование тафономии для вывода различий в мягких тканях между таксонами: пример с использованием базальных и производных форм птерозавров Солнхофена». Наука о природе. 104 (7–8): 65. Bibcode:2017SciNa.104 ... 65B. Дои:10.1007 / s00114-017-1486-0. PMID 28721556.
- ^ Сьюз, Х.-Д. (2009). "Археоптерикс: Der Urvogel von Solnhofen, Петер Веллнхофер ». Журнал палеонтологии позвоночных. 29 (1): 310–311. Дои:10.1080/02724634.2009.10010387.
- ^ Беннет, С.С. (1995). "Статистическое исследование Рамфоринх из Зольнхофенского известняка в Германии: годичные классы одного крупного вида ». Журнал палеонтологии. 69 (3): 569–580. Дои:10.1017 / S0022336000034946.
- ^ Tennant, J.P .; Mannion, P.D .; Апчерч, П. (2016). «Эволюционные отношения и систематика Atoposauridae (Crocodylomorpha: Neosuchia): последствия для возникновения Eusuchia». Зоологический журнал Линнеевского общества. 177 (4): 854–936. Дои:10.1111 / zoj.12400.
- ^ Эберт, М. (2012). «Ракообразные как добыча рыб архипелага Солнхофен». Археоптерикс. 30: 1–4.
- ^ Арратиа, Г. (2016). «Новые замечательные позднеюрские костистые кости из южной Германии: Ascalaboidae n. Fam., Его содержание, морфология и филогенетические отношения». Окаменелости. 19 (1): 31–59. Дои:10.5194 / fr-19-31-2016.
- ^ Эберт, М .; Кёльбл-Эберт, М. (2010). «Морфология передних краев хвостового и других непарных плавников у позднеюрских Aspidorhynchidae и некоторых других актиноптеригийских рыб в свете их функции». Археоптерикс. 28: 1–22.
- ^ Эберт, М. (2014). "Род Белоностомус Agassiz, 1834 (Neopterygii, Aspidorhynchiformes) в поздней юре архипелага Солнхофен с акцентом на Белоностомус кочии Мюнстер, 1836 г. из Эттлинга (Германия) ". Археоптерикс. 32: 15–43.
- ^ Lambers, P.H. (1994). "Галекоморфные рыбы Катур и Амблисемиус в литографическом известняке Солнхофена (Титон), Бавария ». Geobios. 27: 91–99. Дои:10.1016 / S0016-6995 (94) 80024-3.
- ^ Kriwet, J .; Шмитц, Л. (2005). "Новое понимание распространения и палеобиологии рыб-пикнодонтов. Gyrodus". Acta Palaeontologica Polonica. 50 (1).
- ^ Эберт, М .; Lane, J.A .; Кёльбл-Эберт, М. (2016). "Palaeomacrosemius thiollieri, ген. et sp. nov., новый Macrosemiidae (Neopterygii) из верхней юры архипелага Солнхофен (Германия) и Cerin (Франция), с ревизией рода Макросемиус". Журнал палеонтологии позвоночных. 36 (5): e1196081. Дои:10.1080/02724634.2016.1196081.
- ^ Эберт, М. (2012). "Histionotus (Actinopterygii, Macrosemiidae) - Eine Gattung mit vielen Fragezeichen " [Histionotus (Actinopterygii, Macrosemiidae) - род с множеством вопросительных знаков]. Археоптерикс. 30: 5–15.
- ^ Эберт, М. (2019). "Zandtfuro и Schernfeldfuro, Новые роды Halecomorphi (Actinopterygii) из верхнеюрского архипелага Солнхофен ». Журнал палеонтологии позвоночных. 39 (2): e1592759. Дои:10.1080/02724634.2019.1592759.
- ^ Thies, D .; Лейднер, А. (2011). «Акулы и гитаристы (Elasmobranchii) из поздней юры Европы» (PDF). Палеоразнообразие. 4: 63–184.
- ^ Fuchs, D .; Heyng, A.M .; Кеупп, Х. (2013). "Acanthoteuthis problematica Naef, 1922, почти забытый таксон и его роль в интерпретации арматур головоногих моллюсков ». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 269 (3): 241–250. Дои:10.1127/0077-7749/2013/0347.
- ^ Engeser, T .; Рейтнер, Дж. (1992). "Ein neues Exemplar von Belemnoteuthis mayri Engeser & Reitner, 1981 (Coleoidea, Cephalopoda) aus dem Solnhofener Plattenkalk (Untertithonium) von Wintershof, Bayern " [Новый пример Belemnoteuthis mayri Engeser & Reitner, 1981 (Coleoidea, Cephalopoda) из известняков Solnhofen (нижний титон) из Wintershof, Бавария] (PDF). Археоптерикс. 10: 13–17.
- ^ Klug, C .; Fuchs, D .; Schweigert, G .; Röper, M .; Тишлингер, Х. (2015). "Новая анатомическая информация о руках и ластах исключительно сохраненных Плезиотеутис (Coleoidea) из поздней юры Германии ». Швейцарский журнал палеонтологии. 134 (2): 245–255. Дои:10.1007 / s13358-015-0093-у.
- ^ Hess, H .; Эттер, В. (2011). "Жизнь и смерть Saccocoma tenella (Гольдфусс) ". Швейцарский журнал наук о Земле. 104 (1): 99–106. Дои:10.1007 / s00015-011-0059-z.
- ^ Винклер, Н. (2017). «Две новые креветки-пенеиды (Crustacea: Decapoda: Dendrobranchiata) из литографических известняков Солнхофена (верхняя юра, юг Германии)». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 283 (1): 9–24. Дои:10.1127 / njgpa / 2017/0623.
- ^ Lomax, D.R .; Racay, C.A. (2012). "Длинная мортичная тропа Mesolimulus walchi из верхнеюрского литографического известняка Зольнхофена близ Винтерсхофа, Германия ». Ичнос. 19 (3): 175–183. Дои:10.1080/10420940.2012.702704.
- ^ Гёлих, У.Б. (2017). «Каталог коллекций окаменелых птиц Баварской государственной коллекции палеонтологии и геологии в Мюнхене» (PDF). Циттелиана. 89: 331–349.
- ^ Видович, С.У .; Мартил, Д. (2017). "Таксономия и филогения Diopecephalus kochi (Вагнер, 1837) и "Germanodactylus rhamphastinus"(Вагнер, 1851)" (PDF). Геологическое общество, Лондон, Специальные публикации. 455 (1): 125–147. Дои:10.1144 / sp455.12.
- ^ а б c Carannante, G .; Синьор, М .; Вигорито, М. (2006). «Платтенкалк, богатый позвоночными, Пьетрароя (нижний мел, Южные Апеннины, Италия): новая модель». Фации. 52 (4): 555–577. Дои:10.1007 / s10347-006-0075-z.
- ^ а б c Bravi, S .; Гарассино, А (1998). «Новые биостратографические и палеоэкологические наблюдения на« Платтенкалке »нижнего мела (альб) Пьетрарои (Беневенто, Южная Италия) и его скоплении десятиногих ракообразных». Atti Societá Italiana Scienze Naturali, Милан. 138: 119–171.
- ^ Evans, S.E .; Raia, P .; Барбера, К. (2006). "Род ящериц нижнего мела Хометокадмон из Италии". Меловые исследования. 27 (5): 673–683. Дои:10.1016 / j.cretres.2006.03.004.
- ^ Cau, A .; Baiano, M.A .; Райя, П. (2014). «Новый сфенодонтиан (Reptilia, Lepidosauria) из нижнего мела Южной Италии и филогенетическое родство ринхоцефалов Pietraroia Plattenkalk». Меловые исследования. 49: 172–180. Дои:10.1016 / j.cretres.2014.02.001.
- ^ Dal Sasso, C .; Синьор М. (1998). «Исключительная сохранность мягких тканей динозавра теропод из Италии». Природа. 392 (6674): 383–387. Bibcode:1998Натура.392..383D. Дои:10.1038/32884.
- ^ Buscalioni, A.D .; Piras, P .; Vullo, R .; Синьор, М .; Барбера, К. (2011). «Ранняя Eusuchia crocodylomorpha из богатого позвоночными Plattenkalk в Пьетрарои (нижний альб, южные Апеннины, Италия)». Зоологический журнал Линнеевского общества. 163 (S1): S199 – S227. Дои:10.1111 / j.1096-3642.2011.00718.x.
- ^ McGowan, G .; Эванс, С. (1995). «Амфибии Albanerpetontid из мелового периода Испании». Природа. 373 (6510): 143–145. Bibcode:1995Натура 373..143М. Дои:10.1038 / 373143a0.
- ^ Taverne, L .; Capasso, L .; Дель Ре, М. (2020). «Остеология и филогенетические взаимоотношения Gregoriopycnodus bassanii ген. nov., рыба-пикнодонт (Pycnodontidae) из морского альба (нижний мел) Пьетрароя (юг Италии) " (PDF). Гео Эко Троп. 44 (1): 161–174.
- ^ а б Dalla Vecchia, F .; Barbera, C .; Bizzarini, F .; Bravi, S .; Дельфино, М .; Giusberti, L .; Guidotti, G .; Mietto, P .; Palazzoni, C .; Roghi, G .; Синьор, М .; Симона, О. (2005). "Il Cretaceo Marino" [Морской мел]. В Bonfiglio, L. (ред.). Paleontologia dei Vertebrati в Италии: Evoluzione Biologica, Significato Ambientale e Paleogeografico [Палеонтология позвоночных в Италии: биологическая эволюция, экологическое и палеогеографическое значение]. Sezione Scienze della Terra. 6. Memorie del Museo Civico di Storia Naturale di Verona. С. 101–112.
- ^ Taverne, L .; Капассо, Л. (2019). «Остеология Cavinichthys pachylepis ген. и sp. ноя (Teleostei, Crossognathiformes, Pachyrhizodontidae) из Пьетрароя (нижний мел, Кампания, юг Италии) ». Талассия Салентина. 41: 53–70. Дои:10.1285 / i15910725v41p53.
- ^ Taverne, L .; Капассо, Л. (2017). «Остеология и взаимоотношения Caeus («Чанос») Леопольди (Teleostei, Gonorynchiformes, Chanidae) из морского альба (ранний мел) Пьетрароха (Кампания, юг Италии) ». Bollettino dem Museo Civico di Storia Naturale di Verona, Geologia Paleontologia Preistoria. 41: 3–20. S2CID 52025491.
- ^ Taverne, L.P .; Капассо, Л. (2016). "Пересмотр Ionoscopus petrarojae (Ionoscopiformes, Osteichthyes) из альба Пьетрароха (Кампания, юг Италии) ". Талассия Салентина. 38: 65–80. Дои:10.1285 / i15910725v38p65.
- ^ Taverne, L .; Капассо, Л. (2017). "Italophiopsis derasmoi ген. и sp. ноя (Ionoscopiformes, Italophiopsidae fam. Nov.) Из мелового периода Пьетрароя (Италия) ». Талассия Салентина. 39: 9–24. Дои:10.1285 / i15910725v39p9.