Cis-регуляторный модуль - Cis-regulatory module
Было предложено, чтобы эта статья была слился с Цис-регуляторный элемент. (Обсуждать) Предлагается с июля 2020 года. |
СНГ-регулирующий модуль (CRM) является продолжением ДНК, обычно 100–1000 пар оснований ДНК,[1] где ряд факторы транскрипции может связать и регулировать выражение поблизости гены и регулировать их скорость транскрипции. Они обозначены как СНГ потому что они обычно расположены на той же цепи ДНК, что и гены, которые они контролируют, а не транс, который относится к эффектам на гены, расположенные не на той же цепи или дальше, например на факторы транскрипции.[1] Один СНГ-регуляторный элемент может регулировать несколько генов,[2] и наоборот, один ген может иметь несколько СНГ-регулирующие модули.[3] СНГ-регуляторные модули выполняют свою функцию, интегрируя активные факторы транскрипции и связанные с ними кофакторы в определенное время и в определенном месте в ячейке, где эта информация считывается и выдается результат.[4]
Функция генной регуляции
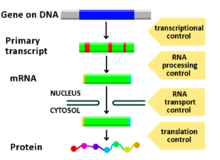
СНГ-регулирующие модули являются одним из нескольких типов функциональных регулирующие элементы. Регуляторные элементы представляют собой сайты связывания факторов транскрипции, которые участвуют в регуляции генов.[1] СНГ-регулирующие модули выполняют большой объем обработки развивающей информации.[1] СНГ-регуляторные модули представляют собой неслучайные кластеры на их указанном целевом сайте, которые содержат сайты связывания факторов транскрипции.[1]
Первоначальное определение представляло цис-регуляторные модули как энхансеры цис-действующей ДНК, которые увеличивали скорость транскрипции из связанных промоутер.[4] Однако это определение было изменено на определение СНГ-регуляторные модули в виде последовательности ДНК с сайтами связывания факторов транскрипции, которые сгруппированы в модульные структуры, включая - но не ограничиваясь ими - области контроля локуса, промоторы, энхансеры, сайленсеры, элементы контроля границ и другие модуляторы.[4]
СНГ-регулирующие модули можно разделить на три класса; усилители, которые положительно регулируют экспрессию генов;[1] изоляторы, которые работают косвенно, взаимодействуя с другими находящимися поблизости СНГ-регулирующие модули; и[1] глушители которые отключают экспрессию генов.[1]
Дизайн СНГ-регуляторные модули таковы, что факторы транскрипции и эпигенетические модификации служат в качестве входных данных, а выходными данными модуля является команда, отдаваемая аппарату транскрипции, который, в свою очередь, определяет скорость транскрипции гена или его включение или выключение.[1] Существует два типа входных сигналов факторов транскрипции: те, которые определяют, когда должен быть экспрессирован целевой ген, и те, которые служат функциональными. водители, которые вступают в силу только в определенных ситуациях во время разработки.[1] Эти входные данные могут поступать из разных временных точек, могут представлять разные сигнальные лиганды или могут исходить из разных доменов или клонов клеток. Однако многое еще остается неизвестным.
Кроме того, регуляция структуры хроматина и ядерной организации также играет роль в определении и контроле функции цис-регуляторных модулей.[4] Таким образом, функции регуляции генов (GRF) обеспечивают уникальную характеристику цис-регуляторного модуля (CRM), связывая концентрации факторов транскрипции (вход) с активностью промотора (выход). Задача состоит в том, чтобы предсказать GRF. Эта проблема до сих пор остается нерешенной. Как правило, функции генной регуляции не используют Логическая логика,[2] хотя в некоторых случаях приближение Логическая логика все еще очень полезно.
Предположение булевой логики
В рамках предположения булевой логики принципы, регулирующие работу этих модулей, включают в себя дизайн модуля, который определяет регулирующую функцию. Что касается разработки, эти модули могут генерировать как положительные, так и отрицательные результаты. Выходные данные каждого модуля являются результатом различных операций, выполняемых над ним. Общие операции включают "ИЛИ" логический вентиль - Этот дизайн указывает, что in выход будет дан, когда задан любой вход [3]. Логический вентиль «И» - в этой конструкции необходимы два разных регулирующих фактора, чтобы гарантировать положительный результат на выходе.[1] «Тумблеры» - эта конструкция возникает, когда сигнальный лиганд отсутствует, а фактор транскрипции присутствует; этот фактор транскрипции действует как доминирующий репрессор. Однако, как только сигнальный лиганд присутствует, роль фактора транскрипции как репрессора устраняется, и может происходить транскрипция.[1]
Другой Логическая логика также могут происходить операции, такие как специфичные для последовательности репрессоры транскрипции, которые, когда они связываются с СНГ-регулирующий модуль приводит к выходу нуля. Кроме того, помимо влияния различных логических операций, на выход модуля регулирования «цис» также будут влиять предшествующие события.[1]4) СНГ-регуляторные модули должны взаимодействовать с другими регулирующими элементами. По большей части, даже при наличии функционального перекрытия между СНГ-регулирующие модули гена, входы и выходы модулей обычно не совпадают.[1]
Хотя предположение о булевой логике важно для системная биология, подробные исследования показывают, что в целом логика регуляции генов не является булевой.[2] Это означает, например, что в случае СНГ-регуляторный модуль регулируется двумя факторами транскрипции, экспериментально определенные функции регуляции генов не могут быть описаны с помощью 16 возможных булевых функций двух переменных. Для исправления этой проблемы были предложены небулевы расширения логики регуляции генов.[2]
Идентификация и вычислительное предсказание
Помимо экспериментального определения CRM, существуют различные биоинформатика алгоритмы их прогнозирования. Большинство алгоритмов пытаются найти значимые комбинации сайтов связывания факторов транскрипции (Сайты связывания ДНК ) в промоутер последовательности коэкспрессируемых генов.[5] Более продвинутые методы сочетают поиск значимых мотивов с корреляцией в экспрессия гена наборы данных между факторы транскрипции и гены-мишени.[6]Оба метода реализованы, например, в ModuleMaster.Другие программы, созданные для выявления и прогнозирования СНГ-регулирующие модули включают:
НАСЕКОМЫЕ 2.0[7] - это веб-сервер, который позволяет осуществлять поиск Цис-регуляторных модулей по всему геному. Программа полагается на определение строгих ограничений среди сайтов связывания факторов транскрипции (TFBS), составляющих модуль, с целью уменьшения количества ложных срабатываний. INSECT разработан так, чтобы быть удобным для пользователя, поскольку он позволяет автоматически извлекать последовательности и несколько визуализаций и ссылок на сторонние инструменты, чтобы помочь пользователям найти те экземпляры, которые с большей вероятностью являются настоящими нормативными сайтами. Алгоритм INSECT 2.0 был ранее опубликован, а алгоритм и теория, лежащие в его основе, объяснены в[8]
Stubb использует скрытые Марковские модели для выявления статистически значимых кластеров комбинаций факторов транскрипции. Он также использует второй родственный геном для повышения точности прогноза модели.[9]
Байесовские сети использовать алгоритм, который объединяет предсказания сайтов и данные тканеспецифической экспрессии для факторов транскрипции и целевых генов, представляющих интерес. В этой модели также используются деревья регрессии, чтобы отобразить взаимосвязь между выявленными СНГ-регуляторный модуль и возможный набор связывающих факторов транскрипции.[10]
CRÈME исследует кластеры сайтов-мишеней на предмет интересующих факторов транскрипции. Эта программа использует базу данных подтвержденных сайтов связывания факторов транскрипции, которые были аннотированы через человеческий геном. Поиск алгоритм применяется к набору данных для идентификации возможных комбинаций факторов транскрипции, которые имеют сайты связывания, близкие к промотору интересующего набора генов. Затем возможные цис-регуляторные модули анализируются статистически, а значимые комбинации представляются графически.[11]
Активный СНГ-регуляторные модули в геномной последовательности трудно идентифицировать. Проблемы с идентификацией возникают из-за того, что часто ученые сталкиваются с небольшим набором известных факторов транскрипции, что затрудняет идентификацию статистически значимых кластеров сайтов связывания факторов транскрипции.[9] Кроме того, высокая стоимость ограничивает использование большого полного генома. мозаичные массивы.[10]
Классификация
СНГ-регуляторные модули могут характеризоваться обработкой информации, которую они кодируют, и организацией их сайтов связывания факторов транскрипции. Кроме того, СНГ-регуляторные модули также характеризуются тем, как они влияют на вероятность, пропорцию и скорость транскрипции.[4]Высокая степень сотрудничества и координации СНГ-регулирующие модули классифицируются как энхансомы.[4] Архитектура и расположение сайтов связывания факторов транскрипции имеют решающее значение, потому что нарушение их расположения может аннулировать функцию.[4]Функциональная гибкость СНГ-регулирующие модули называются рекламными щитами. Их транскрипционный выход является суммирующим эффектом связанных факторов транскрипции.[4]Энхансеры влияют на вероятность активации гена, но практически не влияют на скорость.[4]Модель двоичного ответа действует как переключатель включения / выключения для транскрипции. Эта модель увеличивает или уменьшает количество клеток, транскрибирующих ген, но не влияет на скорость транскрипции.[4]Модель реостатического ответа описывает цис-регуляторные модули как регуляторы скорости инициации транскрипции связанного с ним гена.[4]
Способ действия
СНГ-регуляторные модули могут регулировать свои гены-мишени на больших расстояниях. Было предложено несколько моделей для описания способа, которым эти модули могут связываться с промотором их целевого гена.[4] К ним относятся модель сканирования ДНК, модель петли последовательности ДНК и упрощенная модель отслеживания. В модели сканирования ДНК фактор транскрипции и кофактор сложная форма на СНГ-регуляторный модуль, а затем продолжает двигаться по последовательности ДНК, пока не найдет промотор целевого гена.[4]В петлевой модели фактор транскрипции связывается с СНГ-регулирующий модуль, который затем вызывает зацикливание последовательности ДНК и обеспечивает взаимодействие с промотором целевого гена. Фактор транскрипции -СНГКомплекс -регуляторный модуль вызывает медленное образование петли последовательности ДНК по направлению к промотору-мишени и формирует стабильную петлевую конфигурацию.[4] Модель упрощенного отслеживания объединяет части двух предыдущих моделей.
СНГ-регуляторный модуль в генной регуляторной сети
Функция сеть регуляции генов зависит от архитектуры узлы, функция которого зависит от кратного СНГ-регулирующие модули.[1] Макет СНГ-регуляторные модули могут предоставить достаточно информации для создания пространственных и временных паттернов экспрессии генов.[1] Во время разработки каждый домен, где каждый домен представляет разные пространственные области эмбриона, экспрессии генов будет находиться под контролем разных СНГ-регулирующие модули.[1] Дизайн регулирующих модулей помогает в производстве Обратная связь, кормить вперед, и перекрестные регуляторные петли.[12]
Смотрите также
- Цис-регуляторный элемент
- Энхансосома
- Усилитель
- Оператор
- Оперон
- Промоутер
- Регулирование экспрессии генов
- Трансрегуляторный элемент
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о п q Дэвидсон EH (2006). Регуляторный геном: регуляторные сети генов в развитии и эволюции. Эльзевир. С. 1–86.
- ^ а б c d Тейф В.Б. (2010). «Прогнозирование функций регуляции генов: уроки умеренных бактериофагов». Биофизический журнал. 98 (7): 1247–56. Дои:10.1016 / j.bpj.2009.11.046. ЧВК 2849075. PMID 20371324.
- ^ Бен-Табу де-Леон С., Дэвидсон Э. Х. (2007). «Регулирование генов: сеть контроля генов в разработке» (PDF). Annu Rev Biophys Biomol Struct. 36: 191–212. Дои:10.1146 / annurev.biophys.35.040405.102002. PMID 17291181.
- ^ а б c d е ж грамм час я j k л м п Езёрска Д.М., Джордан К.В., Вэнс К.В. (2009). «Подход системной биологии к пониманию функции цис-регуляторного модуля». Семин. Cell Dev. Биол. 20 (7): 856–862. Дои:10.1016 / j.semcdb.2009.07.007.
- ^ Aerts, S .; и другие. (2003). «Вычислительное обнаружение цис-регуляторных модулей». Биоинформатика. 19 Дополнение 2: ii5–14. Дои:10.1093 / биоинформатика / btg1052. PMID 14534164.
- ^ Wrzodek, Clemens; Шредер, Адриан; Dräger, Андреас; Ванке, Дирк; Berendzen, Kenneth W .; Кронфельд, Марсель; Хартер, Клаус; Зелл, Андреас (2010). «ModuleMaster: новый инструмент для расшифровки сетей регуляции транскрипции». Биосистемы. Ирландия: Эльзевир. 99 (1): 79–81. Дои:10.1016 / j.biosystems.2009.09.005. ISSN 0303-2647. PMID 19819296.
- ^ Парра Р.Г., Рор КО, Койле Д., Перес-Кастро С., Янкилевич П. (2015). «INSECT 2.0: веб-сервер для полногеномного предсказания цис-регуляторных модулей». Биоинформатика. 32 (8): 1229–31. Дои:10.1093 / биоинформатика / btv726. PMID 26656931.
- ^ Рор КО, Парра Р., Янкилевич П., Перес-Кастро С. (2013). «НАСЕКОМЫЕ: IN-silico ПОИСК на наличие сопутствующих факторов транскрипции». Биоинформатика. 29 (22): 2852–8. Дои:10.1093 / биоинформатика / btt506. PMID 24008418.
- ^ а б Синха С., Лян И, Сиггиа Э (2006). «Stubb: программа для обнаружения и анализа цис-регуляторных модулей». Нуклеиновые кислоты Res. 34 (Проблема с веб-сервером): W555 – W559. Дои:10.1093 / нар / gkl224. ЧВК 1538799. PMID 16845069.
- ^ а б Чен Х, Бланшетт М (2007). «Сравнение последовательностей без использования выравниваний: применение к подтипам ВИЧ / SIV». BMC Bioinformatics. 8: 1–17. Дои:10.1186/1471-2105-8-1. ЧВК 1766362. PMID 17199892.
- ^ Шаран Р., Бен-Гур А., Лутс Г. Г., Овчаренко И. (2004). "CREME: Cis-Regulatory Module Explorer для генома человека". Нуклеиновые кислоты Res. 32 (Проблема с веб-сервером): W253 – W256. Дои:10.1093 / нар / гх385. ЧВК 441523. PMID 15215390.
- ^ Ли Э., Дэвидсон Э.Х. (2009). «Построение регуляторных сетей развития генов». Врожденные дефекты Res. 87 (2): 123–130. Дои:10.1002 / bdrc.20152. ЧВК 2747644. PMID 19530131.
внешняя ссылка
- Информация о регуляции генов - вручную составленные списки ресурсов, обзоры, обсуждения в сообществе
- Клеточный дарвинизм
- Регуляция экспрессии генов в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
| Обзор |
| ||||||
|---|---|---|---|---|---|---|---|
| Инженерное дело |
| ||||||
| |||||||