Ornithosuchidae - Ornithosuchidae
| Ornithosuchidae | |
|---|---|
 | |
| Восстановление жизни Орнитосух мусор на ринкозавр Гиперодапедон | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Рептилии |
| Clade: | Псевдозухии |
| Семья: | †Ornithosuchidae фон Хюене, 1908 |
| Типовой вид | |
| †Орнитосухус лесной Ньютон, 1894 г. | |
| Роды | |
†Аэнигмаспина ? | |
Ornithosuchidae является вымерший семья из псевдозухий архозавры (дальние родственники современных крокодилы ) от Триасовый период. Орнитосухиды были четвероногий и факультативно двуногий, что означает, что они могли ходить на двух ногах в течение коротких периодов времени. У них были характерные опущенные морды, уникальные "перевернутые крокодилом" лодыжки и несколько других особенностей, которые палеонтологи используют, чтобы отличить их от других архозавров. Орнитосухиды были географически широко распространены в Карнийский и Norian этапы Поздний триас, с членами, известными из Аргентина, Бразилия, а объединенное Королевство. Четыре рода, Орнитосух, Venaticosuchus, Dynamosuchus,[1] и Riojasuchus в настоящее время известны.[2] Семья была впервые построена немецким палеонтологом. Фридрих фон Хюене в 1908 г.[3]
Описание
Череп
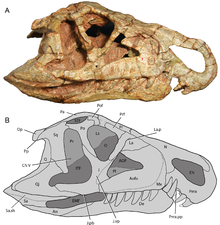
Орнитосухид можно идентифицировать по наличию арочного диастема, щель между зубами в передней части морды. Когда челюсть закрыта, два больших изогнутых зубной (нижняя челюсть) зубы входят в диастему, которая расположена между предчелюстная кость и верхняя челюсть. На стенке диастемы есть два неглубоких углубления для размещения этих зубов. Большие зубные зубы Орнитосух и Riojasuchus располагаются за выпадающим зубом меньшего размера, который выступает из челюсти. Такое положение зубов не наблюдается ни у каких других базальных архозавров. Другой характерной особенностью орнитозухид является их необычно опущенная, нависшая морда, которую можно увидеть у Riojasuchus и Venaticosuchus, но нет Орнитосух.[4]
Несколько других особенностей отличают орнитосухид от всех других ранних архозавров. Орнитосух и Riojasuchus у обоих есть небольшое отверстие, или отверстие между небный и крыловидные кости из нёбо, т. е. нёбо. Контакт между носовой и префронтальные кости черепа небольшой или отсутствует, что исключается большим контактом между лобные и слезные. У других архозавров, в том числе Rauisuchians, этозавры, птерозавры, и динозавроморфы, носо-префронтальный контакт отделяет лобную кость от слезной.[4]
Посткраниальный скелет
Посткраниальный скелет почти полностью известен в Riojasuchus, неполный в Орнитосух, и совершенно неизвестно в Venaticosuchus. В результате неясно, все ли посткраниальные черты, кажущиеся уникальными для орнитозухид, действительно наблюдались у всех членов семьи. Орнитозухиды, известные по приличным посткраниальным останкам, обычно имели около 9 шейный (шея), 14-15 спинной (спина), 3 крестцовый (бедро) и более 20 хвостовых (хвост) позвонки. Над каждым позвонком была пара костных щитков, известных как остеодермы.[2]
В бедренная кость (бедренная кость) имеет ярко выраженный передний вертел. Передний вертел, иногда известный как «малый вертел» (но не имеющий отношения к малый вертел бедренной кости у человека), представляет собой гребень на внешней поверхности бедренной кости, рядом с бедренная головка. Вероятно, это была точка вставки iliofemoralis cranialis мышца, которая помогает поднять ногу. У большинства архозавров и их родственников отсутствует отчетливый передний вертел, но орнитозухиды являются исключением, как и большинство других. динозавроморфы (динозавры и их близкие родственники).[5]
Как и бедренная кость, малоберцовая кость (внешняя кость голени) также имеет отличительную точку для прикрепления мышц. Рассматриваемая мышца - это iliofibularis, который помогает выпрямить конечности. У большинства архозавров iliofibularis вводится в малоберцовую кость с помощью крошечного гребня на проксимальной части малоберцовой кости, около колена. Однако орнитозухиды имеют гораздо более крупную шишковидную форму. iliofibularis точка вставки расположена примерно на полпути вниз по стержню малоберцовой кости. Фитозавры и этозавры также имеют общую точку прикрепления, похожую на шишку, посередине малоберцовой кости, поэтому неясно, является ли случай орнитосухид уникальным случаем конвергентной эволюции или, альтернативно, сохранением черты, независимо утерянной несколькими линиями архозавров.[6]
В отличие от большинства других ранних архозавров, педаль ногтевые (самые дальние кости стоп, образующие когти) сжаты с боков. Они острые и загнутые. Когтевые кости очень глубокие, они выше своей длины, особенно на внутренних пальцах. Этот тип когтей не встречается ни у одного другого триасового архозавра, кроме птерозавров.[4]

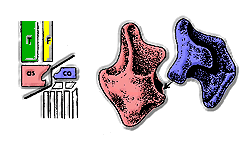
Основные группы архозавров часто отличают друг от друга по строению их лодыжек. У большинства круротарзанов астрагал имеет выпуклый выступ, который вписывается в вогнутое пространство в пяточная кость. Это состояние часто называют «нормальной для крокодила» лодыжкой, поскольку это наиболее распространенный тип лодыжки у круротарзанов. Орнитосухиды уникальны среди круротарсанов и всех других архозавров тем, что у них лодыжка «перевернута крокодилом». В лодыжке, перевернутой крокодилом, вогнутость расположена наоборот: вместо пяточной кости она находится на астрагале. У орнитозухид пяточная кость имеет выпуклый выступ, аналогичный выпуклому выступу у «нормального крокодила» астрагала.[4]
Филогения
Обычно считается, что Ornithosuchidae относятся к более крупным клады Сучия, в которую входят этозавры, рауизухи и крокодиломорфы.[7][8][9] Ниже представлена кладограмма, основанная на работе Nesbitt (2011), показывающая размещение Ornithosuchidae в Archosauriformes.[6]
| Архозаврообразные |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Рекомендации
- ^ а б Мюллер, Родриго Т .; Фон Бачко, М. Белен; Desojo, Julia B .; Несбитт, Стерлинг Дж. (31 января 2020 г.). «Первая орнитосухида из Бразилии и ее макроэволюционные и филогенетические последствия для фаун позднего триаса в Гондване» (PDF). Acta Palaeontologica Polonica. 65. Дои:10.4202 / app.00652.2019.
- ^ а б Бачко, М. Белен фон; Эскурра, Мартин Д. (1 января 2013 г.). "Ornithosuchidae: группа триасовых архозавров с уникальным голеностопным суставом". Геологическое общество, Лондон, Специальные публикации. 379 (1): 187–202. Дои:10.1144 / SP379.4. ISSN 0305-8719.
- ^ Huene, F. von. 1908. "Die Dinosaurier der europäischen Triasformation mit Berücksichtigung der aussereuropäischen Vorkommnisse". Geologische und Paläontologische Abhandlungen 1(Дополнение): 1–419
- ^ а б c d Серено, П. (1991). «Базальные архозавры: филогенетические отношения и функциональные последствия». Журнал палеонтологии позвоночных. 11 (Прил. 4): 1–53. Дои:10.1080/02724634.1991.10011426.
- ^ Langer, Max C .; Бентон, Майкл Дж. (6 ноября 2006 г.). «Ранние динозавры: филогенетическое исследование» (PDF). Журнал систематической палеонтологии. 4 (4): 309–358. Дои:10.1017 / с 1477201906001970. ISSN 1477-2019.
- ^ а б Несбитт, С.Дж. (2011). «Ранняя эволюция архозавров: отношения и происхождение основных клад» (PDF). Бюллетень Американского музея естественной истории. 352: 1–292. Дои:10.1206/352.1. HDL:2246/6112.
- ^ Несбит, S.J .; Норрелл, М.А. (2006). «Чрезвычайная конвергенция в строении тела раннего сициана (Archosauria) и орнитомимидных динозавров (Theropoda)». Труды Королевского общества B. 273 (1590): 1045–1048. Дои:10.1098 / rspb.2005.3426. ЧВК 1560254. PMID 16600879.
- ^ Несбитт, С.Дж. (2007). "Анатомия Effigia okeeffeae (Archosauria, Сучия), тероподоподобная конвергенция и распространение родственных таксонов » (PDF). Бюллетень Американского музея естественной истории. 302: 1–84. Дои:10.1206 / 0003-0090 (2007) 302 [1: TAOEOA] 2.0.CO; 2. HDL:2246/5840.
- ^ Brusatte, S.L .; Benton, M.J .; Desojo, J.B .; Лангер, М. (2010). «Филогения более высокого уровня Archosauria (Tetrapoda: Diapsida)» (PDF). Журнал систематической палеонтологии. 8 (1): 3–47. Дои:10.1080/14772010903537732.
внешняя ссылка
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||

