Мембранный транспорт - Membrane transport
В клеточная биология, мембранный транспорт относится к совокупности механизмов, регулирующих прохождение растворенные вещества Такие как ионы и маленький молекулы через биологические мембраны, которые липидные бислои которые содержат белки встроены в них. Регулирование прохождения через мембрану происходит за счет избирательной проницаемости мембраны - характеристики биологических мембран, которая позволяет им разделять вещества различной химической природы. Другими словами, они могут быть проницаемыми для одних веществ, но не для других.[1]
Движение большинства растворенных веществ через мембрану опосредуется мембранные транспортные белки которые в разной степени специализируются на транспорте определенных молекул. Как разнообразие и физиология различных клетки тесно связано с их способностью привлекать различные внешние элементы, предполагается, что существует группа определенных транспортных белков для каждого типа клеток и для каждой конкретной физиологической стадии[1]. Это дифференциальное выражение регулируемый через дифференциал транскрипция из гены кодирование этих белков и его трансляция, например, через генетико-молекулярные механизмы, но также и на уровне клеточной биологии: производство этих белков может быть активировано клеточные сигнальные пути, на биохимический уровне, или даже находясь в цитоплазматический везикулы.[2]
Фон
Термодинамически поток веществ из одного отсека в другой может происходить в направлении концентрация или же электрохимический градиент или против. Если обмен веществ происходит в направлении градиента, то есть в направлении уменьшения потенциала, нет необходимости во вводе энергии извне системы; Однако если транспорт идет против градиента, он потребует ввода энергии, в данном случае метаболической энергии.[3] Например, классический химический механизм разделения, не требующий добавления внешней энергии, - это диализ. В этой системе полупроницаемая мембрана разделяет два раствора с разной концентрацией одного и того же растворенного вещества. Если мембрана пропускает воду, но не растворенное вещество, вода будет перемещаться в отсек с наибольшей концентрацией растворенного вещества, чтобы установить равновесие в котором энергия системы минимальна. Это происходит из-за того, что вода перемещается от высокой концентрации растворителя к низкой (с точки зрения растворенного вещества происходит обратное), и поскольку вода движется по градиенту, нет необходимости во внешнем подводе энергии.

1. фосфолипид 2. холестерин 3. гликолипид 4. сахар 5. политопный белок (трансмембранный белок) 6. монотопный белок (в данном случае гликопротеин) 7. монотопный белок, заякоренный фосфолипидом 8. периферический монотопный белок (здесь гликопротеин).
Природа биологических мембран, особенно липидов, амфифильный, поскольку они образуют бислои, содержащие внутренний гидрофобный слой и внешний гидрофильный слой. Эта конструкция делает возможной транспортировку простым или пассивная диффузия, который состоит из распространение веществ через мембрану без затрат метаболической энергии и без помощи транспортных белков. Если транспортируемое вещество имеет сетку электрический заряд, он будет двигаться не только в ответ на градиент концентрации, но и на электрохимический градиент из-за мембранный потенциал.
| Тип вещества | Примеры | Поведение |
|---|---|---|
| Газы | CO2, N2, O2 | Проницаемый |
| Маленькие незаряженные полярные молекулы | Мочевина, воды, этиловый спирт | Проницаемый, полностью или частично |
| Большие незаряженные полярные молекулы | глюкоза, фруктоза | Не проницаемый |
| Ионы | K+, Na+, Cl−, HCO3− | Не проницаемый |
| Заряженные полярные молекулы | АТФ, аминокислоты, глюкозо-6-фосфат | Не проницаемый |
Поскольку немногие молекулы способны диффундировать через липидную мембрану, в большинстве транспортных процессов участвуют транспортные белки. Эти трансмембранные белки обладают большим количеством альфа спирали погружен в липидную матрицу. У бактерий эти белки присутствуют в бета-ламина форма.[4] Эта структура, вероятно, включает канал через гидрофильную белковую среду, которая вызывает нарушение высокогидрофобной среды, образованной липидами.[1] Эти белки могут участвовать в транспорте разными способами: они действуют как насосы, приводимые в действие АТФ, то есть метаболической энергией или как каналы облегченной диффузии.
Термодинамика
Физиологический процесс может иметь место только в том случае, если он соответствует основным термодинамический принципы. Мембранный транспорт подчиняется физическим законам, которые определяют его возможности и, следовательно, его биологическую полезность.
Общий принцип термодинамики, который регулирует перенос веществ через мембраны и другие поверхности, заключается в том, что обмен веществ свободная энергия, Δграмм, для перевозки крот вещества с концентрацией C1 в отсеке в другой отсек, где он присутствует на C2 является:[5]

Когда C2 меньше C1, Δграмм отрицательный, и процесс термодинамически благоприятен. Поскольку энергия передается из одного отсека в другой, за исключением случаев вмешательства других факторов, равновесие будет достигнуто где C2= C1, и где Δграмм = 0. Однако есть три обстоятельства, при которых это равновесие не будет достигнуто, обстоятельства, которые жизненно важны для in vivo функционирование биологических мембран:[5]
- Макромолекулы на одной стороне мембраны могут предпочтительно связываться с определенным компонентом мембраны или химически модифицировать его. Таким образом, хотя концентрация растворенного вещества может фактически быть разной на обеих сторонах мембраны, доступность растворенного вещества снижается в одном из отсеков до такой степени, что для практических целей не существует градиента для движения транспорта.
- А электрический потенциал мембраны могут существовать, которые могут влиять на распределение ионов. Например, для переноса ионов снаружи внутрь возможно, что:

Где F Постоянная Фарадея и Δп мембранный потенциал в вольт. Если Δп отрицательно, а Z положительно, вклад члена ZFΔP к Δграмм будет отрицательным, то есть будет способствовать переносу катионов изнутри клетки. Итак, если сохраняется разность потенциалов, состояние равновесия Δграмм = 0 не будет соответствовать эквимолярной концентрации ионов по обе стороны мембраны.
- Если процесс с отрицательным Δграмм связан с транспортным процессом, то глобальная Δграмм будет изменен. Эта ситуация типична для активного транспорта и описывается так:

Где Δграммб соответствует благоприятной термодинамической реакции, такой как гидролиз АТФ, или совместный транспорт соединения, которое перемещается в направлении его градиента.
Виды транспорта
Пассивная диффузия и активная диффузия
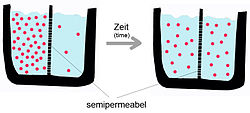
Как упоминалось выше, пассивная диффузия - это спонтанное явление, которое увеличивает энтропия системы и уменьшает свободную энергию.[5] На транспортный процесс влияют характеристики транспортного вещества и природа бислоя. Скорость диффузии чистой фосфолипидной мембраны будет зависеть от:
- градиент концентрации,
- гидрофобность,
- размер,
- заряд, если у молекулы есть чистый заряд.
- Температура
Активный и совместный транспорт
При активном переносе растворенное вещество движется против концентрации или электрохимического градиента; при этом задействованные транспортные белки потребляют метаболическую энергию, обычно АТФ. В первичный активный транспорт гидролиз источника энергии (например, АТФ) происходит непосредственно для транспортировки рассматриваемого растворенного вещества, например, когда транспортные белки АТФаза ферменты. Если гидролиз поставщика энергии является косвенным, как в случае вторичный активный транспорт, используется энергия, запасенная в электрохимическом градиенте. Например, в совместный транспорт используются градиенты определенных растворенных веществ для переноса целевого соединения против его градиента, вызывая рассеивание градиента растворенных веществ. Может показаться, что в этом примере энергия не используется, но требуется гидролиз источника энергии для установления градиента растворенного вещества, переносимого вместе с целевым соединением. Градиент совместно транспортируемый растворенное вещество будет генерироваться за счет использования определенных типов белков, называемых биохимические насосы.[2]
Открытие существования этого типа белка-переносчика произошло в результате изучения кинетики межмембранного транспорта молекул. Для некоторых растворенных веществ было отмечено, что скорость переноса достигла плато при определенной концентрации, выше которой не наблюдалось значительного увеличения скорости поглощения, что указывает на кривая журнала введите ответ. Это было истолковано как свидетельство того, что транспорт опосредованный путем образования комплекса субстрат-переносчик, который концептуально аналогичен комплексу фермент-субстрат кинетика ферментов. Следовательно, каждый транспортный белок имеет константу сродства к растворенному веществу, которая равна концентрации растворенного вещества, когда скорость переноса составляет половину его максимального значения. В случае фермента это эквивалентно Константа Михаэлиса-Ментен.[6]
Некоторыми важными особенностями активного транспорта, помимо его способности вмешиваться даже против градиента, его кинетики и использования АТФ, являются его высокая селективность и простота селективного фармакологического ингибирования.[6]
Вторичные активные белки-переносчики
Вторичные активные белки-переносчики перемещают две молекулы одновременно: одна против градиента, а другая - с его градиентом. Их различают по направленности двух молекул:
- антипортер (также называемый обменником или контр-переносчиком): перемещать молекулу против ее градиента и в то же время перемещать один или несколько ионов по ее градиенту. Молекулы движутся в противоположных направлениях.
- сторонник: перемещать молекулу против ее градиента, перемещая один или несколько различных ионов по их градиенту. Молекулы движутся в одном направлении.
Оба могут быть названы совместные перевозчики.
Насосы

Насос - это белок, который гидролизует АТФ, чтобы транспортировать конкретное растворенное вещество через мембрану, создавая при этом электрохимический градиент. мембранный потенциал. Этот градиент представляет интерес как индикатор состояния ячейки с помощью таких параметров, как Потенциал Нернста. С точки зрения мембранного транспорта градиент представляет интерес, поскольку он способствует снижению энтропии системы в совместный транспорт веществ против их градиента. Одним из наиболее важных насосов в клетках животных является натрий-калиевый насос, который работает через следующий механизм:[7]
- связывание трех Na+ ионы в свои активные центры на помпе, которые связаны с АТФ.
- АТФ гидролизуется, что приводит к фосфорилированию цитоплазматической стороны насоса, что вызывает изменение структуры белка. Фосфорилирование вызывается переносом концевой группы АТФ на остаток аспартат в транспортном белке и последующем высвобождении АДФ.
- изменение структуры в насосе обнажает Na+ к экстерьеру. Фосфорилированная форма насоса имеет низкое сродство к Na+ ионы, поэтому они высвобождаются.
- однажды Na+ освобождаются, насос связывает две молекулы K+ к их соответствующим сайтам связывания на внеклеточной поверхности транспортного белка. Это вызывает дефосфорилирование насоса, возвращая его в его предыдущее конформационное состояние, транспортируя K+ ионы в клетку.
- Нефосфорилированная форма насоса имеет более высокое сродство к Na+ ионов, чем K+ ионы, поэтому два связанных K+ ионы попадают в цитозоль. Связывается АТФ, и процесс начинается снова.
Мембранная селективность
Поскольку основной характеристикой переноса через биологическую мембрану является ее избирательность и ее последующее поведение в качестве барьера для определенных веществ, физиология, лежащая в основе этого явления, была тщательно изучена. Исследования селективности мембран классически подразделяются на исследования, связанные с электролиты и неэлектролиты.
Селективность электролита
Ионные каналы определяют внутренний диаметр, который позволяет прохождение небольших ионов, что связано с различными характеристиками ионов, которые потенциально могут переноситься. Поскольку размер иона связан с его химическим составом, можно предположить, что априори что канал, диаметр поры которого был достаточен для прохождения одного иона, также допускал бы передачу других ионов меньшего размера, однако в большинстве случаев этого не происходит. Помимо размера, существуют две характеристики, которые важны для определения селективности пор мембраны: обезвоживание и взаимодействие иона с внутренними зарядами поры.[6]
Чтобы ион мог пройти через пору, он должен отделиться от молекул воды, которые покрывают его последовательными слоями. сольватация. Склонность к дегидратации или возможность сделать это связаны с размером иона: более крупные ионы могут делать это легче, чем более мелкие, так что пора со слабыми полярными центрами будет предпочтительно пропускать более крупные ионы через ионную поверхность. поменьше.[6]Когда внутренняя часть канала состоит из полярных групп боковых цепей составляющих аминокислот,[7] взаимодействие дегидратированного иона с этими центрами может быть более важным, чем возможность дегидратации, для придания специфичности канала. Например, канал, состоящий из гистидинов и аргининов с положительно заряженными группами, будет избирательно отталкивать ионы той же полярности, но будет способствовать прохождению отрицательно заряженных ионов. Кроме того, в этом случае самые маленькие ионы смогут более тесно взаимодействовать из-за пространственного расположения молекулы (стерильности), что значительно увеличивает заряд-зарядовые взаимодействия и, следовательно, преувеличивает эффект.[6]
Неэлектролитная селективность
Неэлектролиты, вещества, которые обычно являются гидрофобными и липофильными, обычно проходят через мембрану путем растворения в липидном бислое и, следовательно, путем пассивной диффузии. Для тех неэлектролитов, перенос которых через мембрану опосредованный транспортным белком способность к диффузии, как правило, зависит от коэффициент распределения K. Частично заряженные неэлектролиты, которые более или менее полярны, такие как этанол, метанол или мочевина, могут проходить через мембрану через водные каналы, погруженные в мембрану. Не существует эффективного механизма регуляции, ограничивающего этот транспорт, что указывает на внутреннюю уязвимость клеток к проникновению этих молекул.[6]
Создание мембранных транспортных белков
Существует несколько баз данных, которые пытаются построить филогенетические деревья, детализирующие создание белков-переносчиков. Одним из таких ресурсов является База данных классификации транспортеров [8]
Смотрите также
Рекомендации
- ^ а б Лодиш; и другие. (2005). Biología celular y молекулярная (Буэнос-Айрес: изд. Médica Panamericana). ISBN 950-06-1374-3.
- ^ а б Альбертс; и другие. (2004). Молекулярная биология целюлы (Барселона: изд. Омега). ISBN 84-282-1351-8.
- ^ Кромер, А.Х. (1996). Física para ciencias de la vida (на испанском языке) (Reverté ediciones ed.). ISBN 84-291-1808-Х.
- ^ Прескотт, Л. М. (1999). Microbiología (McGraw-Hill Interamericana de España, изд. S.A.U.). ISBN 84-486-0261-7.
- ^ а б c Мэтьюз К. К .; Ван Холд, К.Е .; Ахерн, К.Г. (2003). Bioquímica (3-е изд.). ISBN 84-7829-053-2.
- ^ а б c d е ж Randall D; Burggren, W .; Френч, К. (1998). Eckert Fisiología животное (4-е изд.). ISBN 84-486-0200-5.
- ^ а б Ленингер, Альберт (1993). Принципы биохимии, 2-е изд. (Издательство Worth Publishers). ISBN 0-87901-711-2.
- ^ «База данных классификации транспортеров». Архивировано из оригинал 3 января 2014 г.. Получено 15 июля 2010.