Птичий метаавулавирус 2 - Avian metaavulavirus 2
Эта статья нужно больше медицинские справки за проверка или слишком сильно полагается на основные источники. (Июнь 2015 г.) |
| Птичий метаавулавирус 2 | |
|---|---|
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Учебный класс: | Monjiviricetes |
| Заказ: | Mononegavirales |
| Семья: | Paramyxoviridae |
| Род: | Метаавулавирус |
| Разновидность: | Птичий метаавулавирус 2 |
| Синонимы | |
Птичий метаавулавирус 2, ранее Птичий парамиксовирус 2, это разновидность вируса, принадлежащая к семейству Paramyxoviridae и род Метаавулавирус. Вирус - это вирус с отрицательной цепью РНК содержащий одночастный геном. Птичий метаавулавирус 2 является одним из девяти видов, принадлежащих к роду Метаавулавирус. Самый распространенный серотип из Avulavirinae серотип 1, причина Болезнь Ньюкасла (Без даты).[нужна цитата ] Птичий метаавулавирус 2 как известно, вызывает заболевание, особенно легкое респираторные инфекции во внутреннем домашняя птица, включая индейки и куры, и имеет много экономических последствий для производство яиц и птицеводство.[5] Впервые вирус был выделен из штамма в Юкайпа, Калифорния в 1956 г. С тех пор во всем мире были выделены другие изоляты вируса.

Классификация вирусов
Птичий метаавулавирус 2 представляет собой вирус с отрицательной смысловой РНК с односторонний геном.[5] Это принадлежит Группа V из Балтиморская система классификации вирусов.
Структура
Все Парамиксовирус вирусные частицы состоят из рибонуклеопротеин ядро и липопротеиновый бислой.[7] Птичьи парамиксовирусы содержат вирусную оболочку, которая инкапсулирует сферический вирусный капсид, содержащий геном негативно-смысловой РНК. Члены Paramyxoviridae семья, в том числе Птичий метаавулавирус 2, следуйте «правилу шести», имея геном, кратный шести. Это позволяет правильно упаковать геном в нуклеокапсид.[8] Структура оболочки характеризуется шиповидными выступами двух вирусных белков, слитого белка (F) и белка гемагглютинин-нейраминидаза (Hn). Однако эти белковые прогнозы могут различаться для девяти серотипов птичьих парамиксовирусов.[8] Считается, что матричный белок (M) образует внутренний слой оболочки и взаимодействует с белками F и Hn на внешней стороне оболочки вируса во время проникновения вируса.[8]
Геном
Птичий метаавулавирус 2 имеет геном РНК с отрицательным смыслом, который при входе в клетку должен быть транскрибирован в мРНК для производства вирусных белков и репликации генома. Это было предположено электронная микроскопия, что геном принимает левостороннюю спиральную структуру.[7] Размер генома изолированного штамма Yucaipa, выделенного от кур в Yucaipa, California в 1956 г.,[9] имеет длину 14 904 нуклеотида и содержит лидерную последовательность из 55 нуклеотидов на 3'-конце и концевую последовательность из 154 нуклеотидов на 5'-конце.[10] Однако другие изолированные штаммы Птичий метаавулавирус 2 были изолированы, которые содержат геномы одинаковой, но разной длины. Птичий метаавулавирус 2 один из самых маленьких в Paramyxoviridae семья.[10] Геном кодирует шесть вирусных белков, которые отделены друг от друга межгенные области длиной от 3 до 23 нуклеотидов.[10] Шесть вирусных генов окружены областями транскрипция сигналы остановки и запуска, которые помогают в транскрипции генома и продукции субгеномных РНК. Вирусные геномы большинства серотипов парамиксовирусов кодируют гены нуклеокапсидного белка (N), фосфопротеина (P), матричного белка (M), гибридного белка (F), белка гемагглютинин-нейраминидазы (Hn) и большого белка полимеразы. (L).[8]
Цикл репликации
Вложение и запись
Общей чертой всех парамиксовирусов является наличие двух прикрепленных к мембране гликопротеины, один необходим для прикрепления вириона, а другой - для слияния.[11] Считается, что белки F и Hn, присутствующие на внешней оболочке вируса, способствуют проникновению вируса в клетку-хозяин. Слитый белок (F) является интегральным мембранным белком, обладающим многими свойствами, аналогичными другим слитным вирусам.[11] и принимает непосредственное участие в слиянии вирусов. Считается, что две области гептадных повторов (HR1) и (HR2) образуют структуру пучка из шести спиралей, которая помогает индуцировать слияние вирусной мембраны с мембраной клетки-хозяина.[12] Белок Hn состоит из стебля и шаровидной головки, которая обладает способностью связываться с рецепторами, которые связываются с рецепторами сиаловой кислоты, расположенными на клетках в верхних дыхательных путях хозяина.[11] Предполагается, что взаимодействия между белками Hn и F вызывают конформационные изменения в соответствующих белках, контролирующих прикрепление и слияние вирусного капсида с мембраной клетки-хозяина.[8]
Репликация вируса и производство вирусного белка

Парамиксовирусы кодируют только одну РНК-полимеразу, и считается, что репликация происходит в цитоплазме клетки без участия или проникновения в ядро клетки-хозяина.[7] Мало что известно о том, как полимераза способствует репликации и транскрипции вирусов и как эти процессы могут регулироваться.[13] Кодируемая полимераза состоит из двух белков: большой субъединицы, ответственной за синтез и кэппирование РНК, и кодируемого белка фосфопротеина (P), который действует как кофактор.[13] Как описано, вирусный геном включает межгенные области между вирусными генами со специфическими сигналами запуска и остановки. Полимераза распознает эти цис-действующие элементы внутри вирусного генома и инициирует транскрипцию.[13] Транскрипция сначала начинается на 3'-конце генома и имеет длину 40-55 нуклеотидов. промоутер регион. Полимераза производит субгеномные РНК путем транскрипции каждого гена, высвобождения продуцируемой РНК и последующего определения местонахождения следующего гена для транскрипции цис-действующими элементами, присутствующими в геноме.[13] Считается, что сигнал, при котором полимераза высвобождает транскрибированную РНК одного гена, содержит Урочище Поли У, что привело к закрытию поли (А) хвост к транскрибированной РНК.[13] В отличие от производства вирусной мРНК для синтеза вирусного белка, при репликации вирусного генома полимераза не запускается и не останавливается на указанных цис-действующих элементах между каждым геном. Вместо этого полимераза удлиняет РНК по всей длине генома. Выше описана одна модель для гипотетической вирусной репликации и продукции белка парамиксовирусов, однако существуют и другие модели, которые предполагают, что полимераза инициирует репликацию и транскрипцию в различных областях генома.[13]
Сборка и выпуск
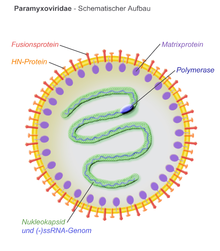
Считается, что матричный белок (M) на внутренней стороне вирусной оболочки играет важную роль в отпочковании вирусоподобных частиц из инфицированной клетки-хозяина.[14] Белок M взаимодействует с белками Hn и F, помогая включить эти белки в вирусные частицы для высвобождения.[8] Была выдвинута гипотеза, что матричные белки парамиксовирусов взаимодействуют с системой сортировки вакуолярных белков хозяина, чтобы инициировать почкование мембраны клетки хозяина. Взаимодействие белков Hn, F и M недостаточно изучено, однако считается, что сборка и высвобождение парамиксовируса зависит от белок-белковых взаимодействий и специфического нацеливания белков на участки клетки-хозяина и мембраны.[14]
Взаимодействие с хозяином
В культуре клеток многие парамиксовирусы экспрессируют цитопатический эффект к синцитий образование, слияние инфицированных клеток с соседними клетками с образованием большой многоядерной клетки.[8] Однако некоторые серотипы птичьего парамиксовируса не показывают образования синцития, а только округляют клетки. Исследования показали, что при инфицировании микроскопические поражения образуются в трахее, легких и, реже, в кишечнике у животных моделей, инфицированных Птичий метаавулавирус 2.[5] Также было показано, что модельные птицы, инфицированные изолятами Птичий метаавулавирус 2 испытывают диарею и показывают увеличенную поджелудочную железу.[5]
Тропизм
Птичий метаавулавирус 2 Считается, что поражает ткани дыхательных путей и в некоторых случаях ткани легких. Белок вирусной оболочки, Hn, связывается с остатками сиаловой кислоты, присутствующими в клетках верхних дыхательных путей. Было обнаружено, что у цыплят у некоторых могут проявляться легкие респираторные симптомы или протекать бессимптомно.[9] Считается, что у индеек наблюдаются более серьезные симптомы, включая синусит и снижение яйценоскости.[9] Сообщалось о других симптомах, включая снижение выводимости и бесплодие яиц.[6]
Распространение вирусов
Птичий метаавулавирус 2 имеет резервуар диких видов птиц и, как известно, вызывает респираторные заболевания и оказывает экономическое влияние на производство яиц и птицеводство[15] Естественные хозяева Птичий метаавулавирус 2 включать индюков и воробьиные. Однако другие хозяева включают цыплят, рельсов и psittacines. Птичий метаавулавирус 2 был зарегистрирован у птиц в Европе, Азии, Северной и Южной Америке. В США было показано, что вирус чаще поражает индеек, чем кур.[9] Изоляты Птичий метаавулавирус 2 были обнаружены в США, Болгарии, Коста-Рике, Африке и многих других местах, что означает Птичий метаавулавирус 2 происходит по всему миру.[5]
Трансмиссия и управление
Передача инфекции происходит медленно между стадами одомашненных индюков в Соединенных Штатах, и способ передачи четко не определен.[6] Предполагается, что инфекции Птичий метаавулавирус 2 у домашней птицы происходят из-за взаимодействия с инфицированными дикими птицами.[6] Профилактические меры могут быть приняты для предотвращения распространения Птичий метаавулавирус 2 в домашних стадах. Такие профилактические меры включают минимизацию взаимодействий между стадами и дикими птицами за счет защиты от птиц участков, где домашняя птица содержится и выращивается, а также надлежащих санитарных методов.[6] Случаев заражения людей не поступало. Птичий метаавулавирус 2[6]
Рекомендации
- ^ Балкема-Бушманн, Анна; и другие. (6 июня 2018 г.). "Реорганизация семьи Paramyxoviridae". Международный комитет по таксономии вирусов (ICTV). Получено 3 мая 2019.
- ^ Rima, Bertus K .; и другие. (Июнь 2016). "Добавление одного (1) нового вида и переименование двенадцати (12) видов в род Авулавирус" (PDF). Международный комитет по таксономии вирусов (ICTV). Получено 3 мая 2019.
Чтобы переименовать следующий таксон (или таксоны): Названия видов в роде Авулавирус. Текущее название Предлагаемое название Птичий парамиксовирус 2 Птичий авулавирус 2
- ^ Третий отчет ICTV Мэтьюз Р. Э. Ф. (1979). Классификация и номенклатура вирусов. Третий отчет Международного комитета по таксономии вирусов. Intervirology 12: 129-296. p217 https://talk.ictvonline.org/ictv/proposals/ICTV%203rd%20Report.pdf
- ^ 1ICTV 5-й отчет Francki, R.I.B., Fauquet, C.M., Knudson, D. L., Brown, F. (ред.) (1991). Классификация и номенклатура вирусов. Пятый отчет Международного комитета по таксономии вирусов. Archives of Virology Supplementum 2, p244. https://talk.ictvonline.org/ictv/proposals/ICTV%205th%20Report.pdf
- ^ а б c d е Варке, Ашвини; Сталлкнехт, Дэвид; Уильямс, Сьюзен М .; Причард, Никки; Мундт, Эгберт (2008). «Сравнительное исследование патогенности и иммуногенности диких птичьих изолятов птичьего парамиксовируса 2, 4 и 6 у кур». Патология птиц. 37 (4): 429–434. Дои:10.1080/03079450802216645. PMID 18622861. S2CID 3995370.
- ^ а б c d е ж «Другие птичьи парамиксовирусные инфекции - Домашняя птица - Ветеринарное руководство Merck». Ветеринарное руководство Merck. Получено 2017-11-02.
- ^ а б c Андерсон, Даниэль (2011). Биология парамиксовирусов. ISBN 9781904455851.
- ^ а б c d е ж грамм Ким, Шин-Хи (2012). "Репликация, нейротропизм и патогенность серотипов птичьего парамиксовируса 1-9 у кур и уток". PLOS ONE. 7 (4): e34927. Bibcode:2012PLoSO ... 734927K. Дои:10.1371 / journal.pone.0034927. ЧВК 3340391. PMID 22558104.
- ^ а б c d Зарьков И.В. (2015). «Обзор инфекции птичьего парамиксовируса серотипа 2 (APMV-2) и первые результаты Болгарии» (PDF).
- ^ а б c Суббья, Мадхури; Сяо, Са; Коллинз, Питер Л .; Самал, Сиба К. (01.10.2008). «Полная последовательность генома птичьего парамиксовируса типа 2 (штамм Yucaipa) и сравнение с другими парамиксовирусами». Вирусные исследования. 137 (1): 40–48. Дои:10.1016 / j.virusres.2008.05.012. ЧВК 2700353. PMID 18603323.
- ^ а б c «Запись парамиксовируса (доступна для скачивания PDF)». ResearchGate. Получено 2017-11-02.
- ^ Ван, Сяо-Цзя; Бай, Я-Дуо; Чжан, Го-Чжун; Чжао, Цзи-Сюнь; Ван, Мин; Гао, Джордж Ф. (апрель 2005 г.). «Исследование структуры и функции пептидов гептадных повторов слитого белка парамиксовируса». Архивы биохимии и биофизики. 436 (2): 316–322. Дои:10.1016 / j.abb.2005.02.004. ЧВК 7094221. PMID 15797244.
- ^ а б c d е ж Noton, Sarah L .; Фирнс, Рэйчел (01.05.2015). «Инициирование и регуляция транскрипции и репликации парамиксовирусов». Вирусология. Выпуск 60-летия. 479-480 (Приложение C): 545–554. Дои:10.1016 / j.virol.2015.01.014. ЧВК 4424093. PMID 25683441.
- ^ а б Пантуа, Гомер Д .; McGinnes, Lori W .; Peeples, Mark E .; Моррисон, Труди Г. (01.11.2006). «Требования к сборке и выпуску частиц, подобных вирусу ньюкаслской болезни». Журнал вирусологии. 80 (22): 11062–11073. Дои:10.1128 / JVI.00726-06. ISSN 0022-538X. ЧВК 1642154. PMID 16971425.
- ^ Александр, Д.Дж. (2000). «Болезнь Ньюкасла и другие птичьи парамиксовирусы» (PDF). Revue Scientifique et Technique (Международное бюро эпизоотии). 19 (2): 443–62. PMID 10935273.