Фототропизм - Phototropism

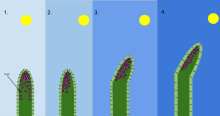






Фототропизм рост организма в ответ на световой раздражитель. Фототропизм чаще всего наблюдается в растения, но также может встречаться в других организмах, таких как грибы. В наиболее удаленных от света клетках растения содержится химическое вещество, называемое ауксин который реагирует, когда возникает фототропизм. Это приводит к тому, что у растения есть удлиненные клетки на самой удаленной от света стороне. Фототропизм - одно из многих растений тропизмы или движения, которые реагируют на внешние раздражители. Рост к источнику света называется положительным фототропизмом, а рост вдали от света - отрицательным фототропизмом (скототропизм).[1] Большинство побегов растений демонстрируют положительный фототропизм и перестраивают свои хлоропласты в листьях, чтобы максимизировать фотосинтетическую энергию и способствовать росту.[2][3] Некоторые кончики побегов лозы проявляют отрицательный фототропизм, который позволяет им расти к темным твердым объектам и взбираться на них. Сочетание фототропизма и гравитропизма позволяет растениям расти в правильном направлении.[4]
Механизм
Есть несколько сигнальных молекул, которые помогают растению определить, откуда исходит источник света, который помогает растению, и это активирует несколько генов, которые изменяют градиенты гормонов, позволяя растению расти в направлении света. Сама верхушка растения известна как колеоптиль, что необходимо при светочувствительности.[2] Средняя часть колеоптиля - это область искривления побега. В Гипотеза Холодного – Вента, разработанный в начале 20-го века, предсказывает, что в присутствии асимметричного света ауксин будет двигаться к затемненной стороне и способствовать удлинению клеток на этой стороне, заставляя растение изгибаться к источнику света.[5] Ауксины активируют протонные насосы, снижая pH в клетках темной стороны растения. Это закисление области клеточной стенки активирует ферменты, известные как экспансины которые разрушают водородные связи в структуре клеточной стенки, делая ее менее жесткой. Кроме того, повышенная активность протонной помпы приводит к тому, что большее количество растворенных веществ попадает в растительные клетки на темной стороне растения, что увеличивает осмотический градиент между симпластом и апопластом этих растительных клеток.[6] Затем вода поступает в клетки по своему осмотическому градиенту, что приводит к увеличению тургорного давления. Снижение прочности клеточной стенки и повышение тургорного давления выше порога текучести[7] заставляет клетки набухать, оказывая механическое давление, которое вызывает фототропное движение.
Вторая группа генов, ШТЫРЬ гены, как было установлено, играют важную роль в фототропизме. Они являются переносчиками ауксина, поэтому считается, что они ответственны за поляризацию ауксина. Конкретно PIN3 был идентифицирован как основной носитель ауксина.[8] Возможно, что фототропины получают свет и ингибируют активность киназы PINOID (PID), которая затем способствует активности PIN3. Эта активация PIN3 приводит к асимметричному распределению ауксина, что затем приводит к асимметричному удлинению клеток в стволе. pin3 мутанты имели более короткие гипокотили и корни, чем у дикого типа, и такой же фенотип наблюдался у растений, выращенных с ингибиторами оттока ауксина.[9] Используя мечение иммуноголотом против PIN3, наблюдали движение белка PIN3. PIN3 обычно локализуется на поверхности гипокотиля и ствола, но также интернализуется в присутствии брефельдина A (BFA), ингибитора экзоцитоза. Этот механизм позволяет изменять положение PIN3 в ответ на раздражитель окружающей среды. Считалось, что белки PIN3 и PIN7 играют роль в индуцированном импульсами фототропизме. Отклики кривизны у мутанта «pin3» были значительно снижены, но лишь немного уменьшены у мутантов «pin7». Между «PIN1», «PIN3» и «PIN7» есть некоторая избыточность, но считается, что PIN3 играет большую роль в индуцированном импульсами фототропизме.[10]
Есть фототропины, которые сильно экспрессируются в верхней части колеоптилей. Есть два основных фототропизма: phot1 и phot2. фото2 одиночные мутанты имеют фототропные ответы, как у дикого типа, но фот1 фот2 двойные мутанты не проявляют фототропных реакций.[4] Суммы ФОТ1 и ФОТ2 присутствуют разные в зависимости от возраста растения и интенсивности света. Есть большое количество ФОТ2 присутствует в зрелом Арабидопсис листья, и это также было замечено в ортологах риса. Выражение ФОТ1 и ФОТ2 меняется в зависимости от наличия синего или красного света. Произошло подавление ФОТ1 мРНК в присутствии света, но активация транскрипта PHOT2. Уровни мРНК и белка, присутствующие в растении, зависели от возраста растения. Это говорит о том, что уровни экспрессии фототропина изменяются с созреванием листьев.[11]Зрелые листья содержат хлоропласты, необходимые для фотосинтеза. Перестройка хлоропластов происходит в различных световых средах, чтобы максимизировать фотосинтез. Есть несколько генов, участвующих в фототропизме растений, включая НПХ1 и NPL1 ген. Оба они участвуют в перестройке хлоропластов.[3] В nph1 и npl1 Было обнаружено, что двойные мутанты обладают сниженными фототропными ответами. Фактически, оба гена являются дублирующими в определении кривизны стебля.
Недавние исследования показали, что в фототропизме растений участвуют несколько киназ AGC, за исключением PHOT1 и PHOT2. Во-первых, PINOID, демонстрируя индуцируемый светом паттерн экспрессии, определяет субклеточное перемещение PIN3 во время фототропных ответов посредством прямого фосфорилирования. Во-вторых, D6PK и его гомологи D6PKL модулируют ауксиновую транспортную активность PIN3, вероятно, также посредством фосфорилирования. В-третьих, перед D6PK / D6PKLs, PDK1.1 и PDK1.2 действуют как важный активатор для этих киназ AGC. Интересно, что разные киназы AGC могут участвовать в разных этапах развития фототропного ответа. D6PK / D6PKL проявляют способность фосфорилировать больше фосфозитов, чем PINOID.
Пять моделей распределения ауксина при фототропизме
В 2012 году Сакаи и Хага[12] обозначил, насколько разные ауксин концентрации могут возникать на затемненной и освещенной стороне стебля, вызывая фототропную реакцию. Пять моделей в отношении корень фототропизм были предложены с использованием Arabidopsis thaliana как исследуемое растение.
- Первая модель
В первой модели входящий свет деактивирует ауксин на светлой стороне растения, позволяя затененной части продолжать рост и в конечном итоге наклонять растение к свету.[12]
- Вторая модель
Во второй модели свет подавляет биосинтез ауксина на светлой стороне растения, тем самым уменьшая концентрацию ауксина по сравнению с незатронутой стороной.[12]
- Третья модель
В третьей модели имеется горизонтальный поток ауксина как со светлой, так и с темной стороны растения. Входящий свет заставляет больше ауксина течь с открытой стороны на затененную сторону, увеличивая концентрацию ауксина на затененной стороне и, таким образом, происходит больший рост.[12]
- Четвертая модель
В четвертой модели показано, как растение получает свет, подавляющий ауксин. базипетальный вниз на открытую сторону, в результате чего ауксин стекает только по затененной стороне.[12]
- Пятая модель
Модель 5 включает в себя элементы как модели 3, так и модели 4. Основной поток ауксина в этой модели идет от верхней части растения вертикально вниз к основанию растения, при этом часть ауксина движется горизонтально от основного потока ауксина к обеим сторонам растения. растение. Получение света подавляет горизонтальный поток ауксина от основного вертикального потока ауксина к облученной открытой стороне. И согласно исследованию Сакаи и Хага, наблюдаемое асимметричное распределение ауксина и последующий фототропный ответ в гипокотилях, по-видимому, наиболее соответствуют этому пятому сценарию.[12]
Влияние длины волны
Фототропизм у таких растений, как Arabidopsis thaliana направляется рецепторами синего света, называемыми фототропины.[13] Другие светочувствительные рецепторы у растений включают: фитохромы это чувство красного света[14] и криптохромы это чувство синего света.[15] Различные органы растения могут проявлять разные фототропные реакции на световые волны разной длины. Кончики стеблей демонстрируют положительную фототропную реакцию на синий свет, а кончики корней - отрицательную фототропную реакцию на синий свет. Как кончики корней, так и большинство кончиков стеблей проявляют положительный фототропизм к красному свету.[нужна цитата ] Криптохромы - это фоторецепторы, которые поглощают синий / УФ-A свет, и они помогают контролировать циркадный ритм растений и время цветения. Фитохромы - это фоторецепторы, которые воспринимают красный / дальний красный свет, но они также поглощают синий свет; они могут контролировать цветение взрослых растений и прорастание семян, среди прочего. Комбинация ответов от фитохромов и криптохромов позволяет растению реагировать на различные виды света.[16] Вместе фитохромы и криптохромы подавляют гравитропизм в гипокотилях и способствуют фототропизму.[2]
Смотрите также
Рекомендации
- ^ Сильный и луч 1975.
- ^ а б c Гоял, А., Сарзинска, Б., Фанкхаузер К. (2012). Фототропизм: на перекрестке свето-сигнальных путей. Ячейка 1-9.
- ^ а б Sakai, T .; Kagawa, T .; Kasahara, M .; Swartz, T.E .; Christie, J.M .; Briggs, W.R .; Wada, M .; Окада, К. (2001). «Arabidopsis nph1 и npl1: рецепторы синего света, которые опосредуют как фототропизм, так и перемещение хлоропластов». PNAS. 98 (12): 6969–6974. Bibcode:2001PNAS ... 98.6969S. Дои:10.1073 / pnas.101137598. ЧВК 34462. PMID 11371609.
- ^ а б Лискум, Э. (2002). Фототропизм: механизмы и результаты. Арабидопсис Книга 1-21.
- ^ Christie, J.M .; Мерфи, А. (2013). «Фототропизм высших растений: новый свет через старые концепции». Американский журнал ботаники. 100 (1): 35–46. Дои:10.3732 / ajb.1200340. PMID 23048016.
- ^ Хагер, Ахим (01.12.2003). «Роль Н + -АТФазы плазматической мембраны в ауксин-индуцированном элонгационном росте: исторические и новые аспекты». Журнал исследований растений. 116 (6): 483–505. Дои:10.1007 / s10265-003-0110-х. ISSN 1618-0860. PMID 12937999. S2CID 23781965.
- ^ Косгроув, Дэниел Дж .; Ван Волкенбург, Элизабет; Клеланд, Роберт Э. (сентябрь 1984 г.). «Релаксация напряжения клеточных стенок и порог текучести для роста: демонстрация и измерение с помощью методов микродавления и психрометра». Planta. 162 (1): 46–54. Дои:10.1007 / BF00397420. ISSN 0032-0935. PMID 11540811. S2CID 6870501.
- ^ Ding, Z .; Galván-Ampudia, C.S .; Demarsy, E .; Langowski, L .; Kleine-Vehn, J .; Fan, Y .; Morita, M.T .; Tasaka, M .; Fankhauser, C .; Offringa, R .; Фримл, Дж. (2011). «Опосредованная светом поляризация переносчика ауксина PIN3 для фототропного ответа у Arabidopsis». Природа клеточной биологии. 13 (4): 447–453. Дои:10.1038 / ncb2208. PMID 21394084. S2CID 25049558.
- ^ Friml, J .; Wisniewska, J .; Бенкова, Е .; Mendgen, K .; Пальме, К. (2002). «Боковое перемещение регулятора оттока ауксина PIN3 опосредует тропизм у Arabidopsis». Природа. 415 (6873): 806–809. Bibcode:2002Натура 415..806F. Дои:10.1038 / 415806a. PMID 11845211. S2CID 4348635.
- ^ Haga, K .; Сакаи, Т. (2012). «Пин-носители оттока ауксина необходимы для импульсного, но не непрерывного светоиндуцированного фототропизма у Arabidopsis». Физиология растений. 160 (2): 763–776. Дои:10.1104 / стр.112.202432. ЧВК 3461554. PMID 22843667.
- ^ Labuz, J .; Штательман, О .; Banas, A. K .; Габрис, Х. (2012). «Экспрессия фототропинов в листьях Arabidopsis: регуляция развития и света». Журнал экспериментальной ботаники. 63 (4): 1763–1771. Дои:10.1093 / jxb / ers061. PMID 22371325.
- ^ а б c d е ж Сакаи, Т; Хага, К. (2012). «Молекулярно-генетический анализ фототропизма арабидопсиса». Физиология растений и клеток. 53 (9): 1517–34. Дои:10,1093 / шт / шт 111. ЧВК 3439871. PMID 22864452.
- ^ «Фототропины: фоторецепторы, обеспечивающие новый фотохимический механизм передачи сигналов». Архивировано из оригинал на 2015-11-18. Получено 2016-04-16.
- ^ «Фитохром». plantphys.info. Получено 2016-04-16.
- ^ Эккардт, Н. А. (1 мая 2003 г.). «Компонент пути передачи сигналов синего света криптохрома». Растительная клетка онлайн. 15 (5): 1051–1052. Дои:10.1105 / tpc.150510. ЧВК 526038.
- ^ Маккошам, С., Поцелуй, Дж. З. (2011). Зеленый свет влияет на фототропизм на основе синего света в гипокотилях Arabidopsis thaliana. Ботаническое общество Торри 138 (4), 409-417.
Библиография
- Сильный, Дональд Р .; Рэй, Томас С. (1 января 1975 г.). «Поведение дерева-хозяина тропической лозы (Монстера гигантская) скототропизмом ». Наука. 190 (4216): 804–806. Bibcode:1975Наука ... 190..804С. Дои:10.1126 / science.190.4216.804. JSTOR 1741614. S2CID 84386403.CS1 maint: ref = harv (связь)
внешняя ссылка
 СМИ, связанные с Фототропизм в Wikimedia Commons
СМИ, связанные с Фототропизм в Wikimedia Commons- Замедленные фильмы, Растения в движении