Функциональная геномика - Functional genomics
Функциональная геномика это область молекулярная биология это пытается описать ген (и белок ) функции и взаимодействия. Функциональная геномика использует обширные данные, генерируемые геномный и транскриптомный проекты (такие как проекты секвенирования генома и Секвенирование РНК ). Функциональная геномика фокусируется на динамических аспектах, таких как ген транскрипция, перевод, регуляция экспрессии генов и белок-белковые взаимодействия, в отличие от статических аспектов геномной информации, таких как Последовательность ДНК или конструкции. Ключевой характеристикой исследований функциональной геномики является их общегеномный подход к этим вопросам, как правило, с использованием высокопроизводительных методов, а не более традиционного подхода «ген за геном».

Определение и цели функциональной геномики
Чтобы понять функциональную геномику, важно сначала определить функцию. В своей статье[1] Graur et al. определить функцию двумя возможными способами. Это «Выбранный эффект» и «Причинная роль». Функция «Выбранный эффект» относится к функции, для которой выбирается признак (ДНК, РНК, белок и т. Д.). Функция «Причинная роль» относится к функции, для которой характеристика достаточна и необходима. Функциональная геномика обычно проверяет определение функции «Причинная роль».
Цель функциональной геномики - понять функцию генов или белков, в конечном итоге всех компонентов генома. Термин функциональная геномика часто используется для обозначения многих технические подходы изучать организм гены и белки, включая «биохимические, клеточные и / или физиологические свойства каждого генного продукта»[2] в то время как некоторые авторы включают исследование негенные элементы в их определении.[3] Функциональная геномика может также включать исследования естественных генетическая вариация через некоторое время (например, развитие организма) или Космос (например, области его тела), а также функциональные нарушения, такие как мутации.
Обещание функциональной геномики состоит в том, чтобы генерировать и синтезировать геномные и протеомные знания для понимания динамических свойств организма. Это потенциально может дать более полную картину того, как геном определяет функцию, по сравнению с исследованиями отдельных генов. Интеграция данных функциональной геномики часто является частью системная биология подходы.
Методы и приложения
Функциональная геномика включает функциональные аспекты самого генома, такие как мутация и полиморфизм (такие как однонуклеотидный полиморфизм (SNP) анализ), а также измерение молекулярной активности. Последние включают в себя ряд "-омики " такие как транскриптомика (экспрессия гена ), протеомика (производство белка ), и метаболомика. Функциональная геномика в основном использует мультиплекс методы измерения численности многих или всех генных продуктов, таких как мРНК или белки в пределах биологический образец. Более сфокусированный подход к функциональной геномике может проверить функцию всех вариантов одного гена и количественно оценить эффекты мутантов, используя секвенирование в качестве считывания активности. Вместе эти методы измерения направлены на количественное определение различных биологических процессов и улучшение нашего понимания функций и взаимодействий генов и белков.
На уровне ДНК
Картирование генетического взаимодействия
Систематическое попарное удаление генов или ингибирование экспрессии генов можно использовать для идентификации генов со связанной функцией, даже если они не взаимодействуют физически. Эпистаз относится к тому факту, что эффекты нокаута двух разных генов могут не складываться; то есть фенотип, который возникает при ингибировании двух генов, может отличаться от суммы эффектов единичных нокаутов.
ДНК / белковые взаимодействия
Белки, образованные в результате трансляции мРНК (информационная РНК, кодированная информация ДНК для синтеза белка), играют важную роль в регулировании экспрессии генов. Чтобы понять, как они регулируют экспрессию генов, необходимо определить последовательности ДНК, с которыми они взаимодействуют. Были разработаны методы определения участков ДНК-белковых взаимодействий. К ним относятся Чип-секвенирование, Секвенирование CUT & RUN и телефонные карточки.[4]
Анализ доступности ДНК
Были разработаны анализы для определения доступных участков генома. Эти области открытого хроматина являются кандидатами в регуляторные области. Эти тесты включают ATAC-seq, ДНКаза-Seq и FAIRE-Seq.
На уровне РНК
Микрочипы

Микроматрицы измеряют количество мРНК в образце, которое соответствует данному гену или последовательности ДНК зонда. Последовательности зондов иммобилизуют на твердой поверхности и дают возможность гибридизировать с флуоресцентно меченой «целевой» мРНК. Интенсивность флуоресценции пятна пропорциональна количеству последовательности-мишени, которая гибридизировалась с этим пятном, и, следовательно, количеству этой последовательности мРНК в образце. Микромассивы позволяют идентифицировать гены-кандидаты, участвующие в данном процессе, на основе различий между уровнями транскриптов для различных условий и общих паттернов экспрессии с генами с известной функцией.
МУДРЕЦ
Серийный анализ экспрессии генов (SAGE) - альтернативный метод анализа, основанный на секвенировании РНК, а не на гибридизации. SAGE полагается на секвенирование тегов из 10–17 пар оснований, уникальных для каждого гена. Эти теги производятся из поли-А мРНК и лигировали встык перед секвенированием. SAGE дает объективное измерение количества транскриптов на клетку, так как оно не зависит от предварительных знаний о том, какие транскрипты изучать (как это делают микроматрицы).
Секвенирование РНК
Как отмечалось в 2016 году, в последние годы секвенирование РНК использовало технологию микрочипов и SAGE и стало наиболее эффективным способом изучения транскрипции и экспрессии генов. Обычно это делается секвенирование следующего поколения.[5]
Подмножество секвенированных РНК - это малые РНК, класс некодирующих молекул РНК, которые являются ключевыми регуляторами транскрипционного и посттранскрипционного подавления генов, или Подавление РНК. Секвенирование нового поколения - это золотой стандарт для некодирующая РНК открытие, профилирование и анализ выражения.
Массивно параллельные репортерные анализы (MPRA)
Массивно-параллельные репортерные анализы - это технология для проверки цис-регуляторной активности последовательностей ДНК.[6][7] В MPRA используется плазмида с синтетическим цис-регуляторным элементом перед промотором, управляющим синтетическим геном, таким как зеленый флуоресцентный белок. Библиотека цис-регуляторных элементов обычно тестируется с использованием MPRA, библиотека может содержать от сотен до тысяч цис-регуляторных элементов. Цис-регуляторная активность элементов оценивается с использованием нижестоящей репортерной активности. Активность всех членов библиотеки анализируется параллельно с использованием штрих-кодов для каждого цис-регуляторного элемента. Одним из ограничений MPRA является то, что активность анализируется на плазмиде и может не улавливать все аспекты регуляции гена, наблюдаемые в геноме.
STARR-seq
STARR-seq - это метод, аналогичный MPRA, для анализа энхансерной активности случайно разрезанных геномных фрагментов. В оригинальной публикации[8] случайно разрезанные фрагменты генома дрозофилы помещали ниже минимального промотора. Кандидаты в энхансеры среди случайно разрезанных фрагментов будут транскрибировать себя с использованием минимального промотора. Используя секвенирование в качестве считывания и контроль вводимых количеств каждой последовательности, этим методом оценивают силу предполагаемых усилителей.
Perturb-seq
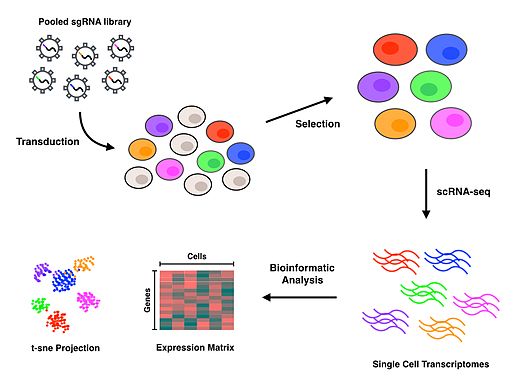
Perturb-seq сочетает опосредованные CRISPR нокдауны генов с экспрессией одноклеточных генов. Линейные модели используются для расчета влияния нокдауна одного гена на экспрессию нескольких генов.
На уровне белка
Дрожжевая двугибридная система
Дрожжи двухгибридный скрининг (Y2H) тестирует белок-«приманку» против многих потенциально взаимодействующих белков («жертва»), чтобы определить физические взаимодействия белок-белок. Эта система основана на факторе транскрипции, первоначально GAL4,[9] чьи отдельные ДНК-связывающие домены и домены активации транскрипции необходимы для того, чтобы белок вызывал транскрипцию репортерного гена. При скрининге Y2H белок «приманка» сливается со связывающим доменом GAL4, и библиотека потенциальных белков «жертвы» (взаимодействующих) рекомбинантно экспрессируется в векторе с доменом активации. Взаимодействие in vivo белков приманки и жертвы в дрожжевой клетке приводит к тому, что активационный и связывающий домены GAL4 достаточно близко друг к другу, чтобы приводить к экспрессии репортерный ген. Также возможно систематически тестировать библиотеку белков-приманок против библиотеки белков-жертв, чтобы определить все возможные взаимодействия в клетке.
AP / MS
Аффинная очистка и масс-спектрометрии (AP / MS) может идентифицировать белки, которые взаимодействуют друг с другом в комплексах. Белковые комплексы могут образовываться вокруг определенного белка-«приманки». Белок приманки идентифицируется с помощью антитела или рекомбинантной метки, что позволяет экстрагировать его вместе с любыми белками, которые образовали с ним комплекс. Затем белки перевариваются в короткие пептид Фрагменты и масс-спектрометрия используются для идентификации белков на основе отношения массы к заряду этих фрагментов.
Глубокое мутационное сканирование
При глубоком мутационном сканировании сначала синтезируются все возможные аминокислотные изменения в данном белке. Активность каждого из этих вариантов белка анализируется параллельно с использованием штрих-кодов для каждого варианта. Сравнивая активность с белком дикого типа, идентифицируют эффект каждой мутации. Хотя возможно проанализировать каждое возможное изменение отдельной аминокислоты из-за комбинаторики, две или более одновременных мутации трудно проверить. Эксперименты по глубокому мутационному сканированию также использовались для определения структуры белка и белок-белковых взаимодействий.
Методы потери функции
Мутагенез
Функцию генов можно исследовать, систематически «выбивая» гены один за другим. Это делается либо удаление или нарушение функции (например, инсерционный мутагенез ), и полученные организмы проверяются на фенотипы, которые дают ключ к разгадке функции нарушенного гена *
РНКи
Методы РНК-интерференции (РНКи) могут использоваться для временного замалчивания или подавления экспрессии генов с использованием двухцепочечной РНК из ~ 20 пар оснований, обычно доставляемой путем трансфекции синтетических ~ 20-мерных коротко интерферирующих молекул РНК (миРНК) или коротких молекул, кодируемых вирусами -шпильчатые РНК (кшРНК). Скрининг РНКи, обычно выполняемый в анализах на основе клеточных культур или экспериментальных организмов (таких как C. elegans) может использоваться для систематического нарушения почти каждого гена в геноме или подмножеств генов (субгеномов); возможные функции нарушенных генов могут быть определены на основе наблюдаемых фенотипы.
CRISPR-экраны

CRISPR-Cas9 был использован для мультиплексной делеции генов в клеточных линиях. Количественная оценка количества направляющих РНК для каждого гена до и после эксперимента может указать на важные гены. Если РНК-проводник разрушает важный ген, это приведет к потере этой клетки и, следовательно, будет истощение этой конкретной РНК-проводника после скрининга. В недавнем эксперименте CRISPR-cas9 на клеточных линиях млекопитающих было обнаружено, что около 2000 генов являются важными во многих клеточных линиях.[11][12] Некоторые из этих генов были важны только для одной клеточной линии. Большинство генов входят в состав мультибелковых комплексов. Этот подход может использоваться для определения синтетической летальности с использованием соответствующего генетического фона. CRISPRi и CRISPRa аналогичным образом позволяют использовать экраны потери функции и усиления функции. CRISPRi идентифицировал ~ 2100 основных генов в клеточной линии K562.[13][14] Скрины делеции CRISPR также использовались для выявления потенциальных регуляторных элементов гена. Например, была опубликована методика под названием ScanDel, в которой использовалась эта попытка. Авторы удалили области за пределами интересующего гена (HPRT1, вовлеченный в менделевское расстройство) в попытке идентифицировать регуляторные элементы этого гена.[15] Gassperini et al. не идентифицировали какие-либо дистальные регуляторные элементы для HPRT1 с использованием этого подхода, однако такие подходы могут быть распространены на другие интересующие гены.
Функциональные аннотации генов
Аннотации генома
Предполагаемые гены могут быть идентифицированы путем сканирования генома на предмет областей, которые могут кодировать белки, на основе таких характеристик, как длина открытые рамки для чтения, последовательности инициации транскрипции и полиаденилирование места. Последовательность, идентифицированная как предполагаемый ген, должна быть подтверждена дополнительными доказательствами, такими как сходство с последовательностями кДНК или EST из того же организма, сходство предсказанной белковой последовательности с известными белками, ассоциация с промоторными последовательностями или доказательство того, что мутация последовательности вызывает наблюдаемый фенотип.
Подход с розеттским камнем
Подход Розеттского камня - это вычислительный метод для предсказания функции белков de novo. Он основан на гипотезе о том, что некоторые белки, участвующие в данном физиологическом процессе, могут существовать как два отдельных гена в одном организме и как один ген в другом. Геномы сканируются на предмет последовательностей, независимых в одном организме и в одной открытой рамке считывания в другом. Если два гена слились, предполагается, что у них есть сходные биологические функции, которые делают такую совместную регуляцию выгодной.
Методы биоинформатики для функциональной геномики
Из-за большого количества данных, полученных с помощью этих методов, и желания найти биологически значимые закономерности, биоинформатика имеет решающее значение для анализа данных функциональной геномики. Примеры методов в этом классе: кластеризация данных или Анализ главных компонентов для неконтролируемых машинное обучение (определение класса), а также искусственные нейронные сети или опорные векторные машины для машинного обучения с учителем (прогнозирование класса, классификация ). Анализ функционального обогащения используется для определения степени избыточной или недостаточной экспрессии (положительные или отрицательные регуляторы в случае скрининга РНКи) функциональных категорий относительно фоновых наборов. Генная онтология анализ обогащения на основе ДЭВИД и анализ обогащения набора генов (GSEA),[16] анализ пути от Ingenuity [17] и студия Pathway[18] и анализ на основе белковых комплексов COMPLEAT.[19]

Были разработаны новые вычислительные методы для понимания результатов эксперимента по глубокому мутационному сканированию. 'phydms' сравнивает результат эксперимента по глубокому мутационному сканированию с филогенетическим деревом.[20] Это позволяет пользователю сделать вывод, накладывает ли процесс отбора в природе аналогичные ограничения на белок, как показывают результаты глубокого мутационного сканирования. Это может позволить экспериментатору выбирать между различными экспериментальными условиями в зависимости от того, насколько хорошо они отражают природу. Глубокое мутационное сканирование также использовалось для вывода белок-белковых взаимодействий.[21] Авторы использовали термодинамическую модель для прогнозирования эффектов мутаций в различных частях димера. Глубокая мутационная структура также может быть использована для вывода структуры белка. Сильный положительный эпистаз между двумя мутациями при глубоком мутационном сканировании может указывать на две части белка, которые расположены близко друг к другу в трехмерном пространстве. Затем эту информацию можно использовать для определения структуры белка. Доказательство принципа этого подхода было продемонстрировано двумя группами, использовавшими белок GB1.[22][23]
Результаты экспериментов MPRA потребовали подходов машинного обучения для интерпретации данных. Модель SVM с пробелом k-mer была использована для вывода kmers, которые обогащены цис-регуляторными последовательностями с высокой активностью по сравнению с последовательностями с более низкой активностью.[24] Эти модели обеспечивают высокую предсказательную силу. Для интерпретации результатов этих многомерных экспериментов также использовались подходы глубокого обучения и случайного леса.[25] Эти модели начинают способствовать лучшему пониманию функции некодирующей ДНК в отношении генной регуляции.
Проекты консорциума, ориентированные на функциональную геномику
Проект ENCODE
Проект ENCODE (Энциклопедия элементов ДНК) - это углубленный анализ генома человека, цель которого - идентифицировать все функциональные элементы геномной ДНК как в кодирующих, так и в некодирующих областях. Важные результаты включают доказательства массивов геномных плиток, что большинство нуклеотидов транскрибируются как кодирующие транскрипты, некодирующие РНК или случайные транскрипты, открытие дополнительных сайтов регуляции транскрипции, дальнейшее выяснение механизмов модификации хроматина.
Проект экспрессии генотипа и ткани (GTEx)

Проект GTEx - это проект в области генетики человека, направленный на понимание роли генетической изменчивости в формировании вариаций транскриптома в разных тканях. В рамках проекта были собраны различные образцы тканей (> 50 различных тканей) от более чем 700 вскрытых доноров. В результате собрано более 11 000 образцов. GTEx помог понять совместное использование тканей и тканевую специфичность EQTLs.[26]
Смотрите также
Рекомендации
- ^ Граур Д., Чжэн Ю., Прайс Н, Азеведо РБ, Зуфалл Р.А., Эльхайк Э. (20 февраля 2013 г.). «О бессмертии телевизоров:« функция »в геноме человека согласно безэволюционному евангелию ENCODE». Геномная биология и эволюция. 5 (3): 578–90. Дои:10.1093 / gbe / evt028. ЧВК 3622293. PMID 23431001.
- ^ Гибсон G, Muse SV. Учебник по геномной науке (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates.
- ^ Певснер Дж (2009). Биоинформатика и функциональная геномика (2-е изд.). Хобокен, Нью-Джерси: Уайли-Блэквелл.
- ^ Ван Х, Мэйхью Д., Чен Х, Джонстон М., Митра Р. Д. (май 2011 г.). «Телефонные карточки позволяют мультиплексную идентификацию геномных мишеней ДНК-связывающих белков». Геномные исследования. 21 (5): 748–55. Дои:10.1101 / гр.114850.110. ЧВК 3083092. PMID 21471402.
- ^ Hrdlickova R, Toloue M, Tian B (январь 2017 г.). «Методы RNA-Seq для анализа транскриптомов». Междисциплинарные обзоры Wiley: РНК. 8 (1): e1364. Дои:10.1002 / wrna.1364. ЧВК 5717752. PMID 27198714.
- ^ Квасниески JC, Fiore C, Chaudhari HG, Cohen BA (октябрь 2014 г.). «Высокопроизводительное функциональное тестирование предсказаний сегментации ENCODE». Геномные исследования. 24 (10): 1595–602. Дои:10.1101 / гр.173518.114. ЧВК 4199366. PMID 25035418.
- ^ Патвардхан Р.П., Хиатт Дж. Б., Виттен Д. М., Ким М. Дж., Смит Р. П., Мэй Д. и др. (Февраль 2012 г.). «Массивно параллельное функциональное вскрытие энхансеров млекопитающих in vivo». Природа Биотехнологии. 30 (3): 265–70. Дои:10.1038 / nbt.2136. ЧВК 3402344. PMID 22371081.
- ^ Арнольд С.Д., Герлах Д., Стельцер С., Борынь М., Рат М., Старк А. (март 2013 г.). «Полногеномные количественные карты активности энхансеров, идентифицированные с помощью STARR-seq». Наука. 339 (6123): 1074–7. Bibcode:2013Научный ... 339.1074А. Дои:10.1126 / science.1232542. PMID 23328393. S2CID 54488955.
- ^ Поля С, Сонг О (июль 1989 г.). «Новая генетическая система для обнаружения белок-белковых взаимодействий». Природа. 340 (6230): 245–6. Bibcode:1989Натура.340..245F. Дои:10.1038 / 340245a0. PMID 2547163. S2CID 4320733.
- ^ Tian S, Muneeruddin K, Choi MY, Tao L, Bhuiyan RH, Ohmi Y, Furukawa K, Furukawa K, Boland S, Shaffer SA, Adam RM, Dong M (27 ноября 2018 г.). «Полногеномный CRISPR-скрининг на токсины шига и рицин выявляет белки Гольджи, критические для гликозилирования». PLOS Биология. 16 (11). e2006951. Дои:10.1371 / journal.pbio.2006951. ЧВК 6258472. PMID 30481169.
- ^ Харт Т., Чандрашекхар М., Ареггер М., Стейнхарт З., Браун К.Р., МакЛауд Дж. И др. (Декабрь 2015 г.). «Скрины CRISPR с высоким разрешением выявляют гены приспособленности и специфические для генотипа функции рака». Клетка. 163 (6): 1515–26. Дои:10.1016 / j.cell.2015.11.015. PMID 26627737.
- ^ Shalem O, Sanjana NE, Hartenian E, Shi X, Scott DA, Mikkelson T. и др. (Январь 2014). «Скрининг нокаута CRISPR-Cas9 в масштабе генома в клетках человека». Наука. 343 (6166): 84–87. Bibcode:2014Наука ... 343 ... 84S. Дои:10.1126 / science.1247005. ЧВК 4089965. PMID 24336571.
- ^ Гилберт Л.А., Хорлбек М.А., Адамсон Б., Вильялта Дж. Э., Чен Ю., Уайтхед Е. Х. и др. (Октябрь 2014 г.). "Контроль репрессии и активации генов, опосредованный CRISPR в масштабе генома". Клетка. 159 (3): 647–61. Дои:10.1016 / j.cell.2014.09.029. ЧВК 4253859. PMID 25307932.
- ^ Хорлбек М.А., Гилберт Л.А., Виллалта Дж. Э., Адамсон Б., Пак Р. А., Чен Ю. и др. (Сентябрь 2016 г.). «Компактные и высокоактивные библиотеки нового поколения для CRISPR-опосредованной репрессии и активации генов». eLife. 5. Дои:10.7554 / eLife.19760. ЧВК 5094855. PMID 27661255.
- ^ Гасперини, Молли; Финдли, Грегори М .; Маккенна, Аарон; Милбанк, Дженнифер Х .; Ли, Чоли; Чжан, Мелисса Д .; Кусанович, Даррен А .; Шендуре, Джей (август 2017 г.). «CRISPR / Cas9-опосредованное сканирование регуляторных элементов, необходимых для экспрессии HPRT1 через тысячи больших запрограммированных геномных делеций». Американский журнал генетики человека. 101 (2): 192–205. Дои:10.1016 / j.ajhg.2017.06.010. ЧВК 5544381. PMID 28712454.
- ^ Субраманиан А., Тамайо П., Мутха В.К., Мукерджи С., Эберт Б.Л., Джиллетт М.А. и др. (Октябрь 2005 г.). «Анализ обогащения набора генов: основанный на знаниях подход к интерпретации профилей экспрессии в масштабе всего генома». Труды Национальной академии наук Соединенных Штатов Америки. 102 (43): 15545–50. Bibcode:2005ПНАС..10215545С. Дои:10.1073 / pnas.0506580102. ЧВК 1239896. PMID 16199517.
- ^ "Системы изобретательности". Архивировано из оригинал на 1999-01-25. Получено 2007-12-31.
- ^ "Геномика Ариадны: Pathway Studio". Архивировано из оригинал на 2007-12-30. Получено 2007-12-31.
- ^ Vinayagam A, Hu Y, Kulkarni M, Roesel C, Sopko R, Mohr SE, Perrimon N (февраль 2013 г.). «Основа анализа на основе белковых комплексов для высокопроизводительных наборов данных». Научная сигнализация. 6 (264): RS5. Дои:10.1126 / scisignal.2003629. ЧВК 3756668. PMID 23443684.
- ^ Хилтон СК, Доуд МБ, Блум ДжейДи (2017). "phydms: программное обеспечение для филогенетического анализа на основе глубокого мутационного сканирования". PeerJ. 5: e3657. Дои:10.7717 / peerj.3657. ЧВК 5541924. PMID 28785526.
- ^ Дисс Дж., Ленер Б. (апрель 2018 г.). «Генетический ландшафт физического взаимодействия». eLife. 7. Дои:10.7554 / eLife.32472. ЧВК 5896888. PMID 29638215.
- ^ Schmiedel, Jörn M .; Ленер, Бен (17 июня 2019 г.). «Определение белковых структур с помощью глубокого мутагенеза». Природа Генетика. 51 (7): 1177–1186. Дои:10.1038 / с41588-019-0431-х. PMID 31209395.
- ^ Роллинз, Натан Дж .; Брок, Келли П .; Poelwijk, Франк Дж .; Стиффлер, Майкл А .; Готье, Николас П .; Сандер, Крис; Маркс, Дебора С. (17 июня 2019 г.). «Выявление трехмерной структуры белка на основе сканирования глубоких мутаций». Природа Генетика. 51 (7): 1170–1176. Дои:10.1038 / s41588-019-0432-9. ЧВК 7295002. PMID 31209393.
- ^ Ганди М., Ли Д., Мохаммад-Нури М., Бир М.А. (июль 2014 г.). «Улучшенное предсказание регуляторной последовательности с использованием функций k-mer с разрывом». PLOS вычислительная биология. 10 (7): e1003711. Bibcode:2014PLSCB..10E3711G. Дои:10.1371 / journal.pcbi.1003711. ЧВК 4102394. PMID 25033408.
- ^ Ли И, Ши В., Вассерман В.В. (май 2018 г.). «Полногеномное предсказание цис-регуляторных регионов с использованием контролируемых методов глубокого обучения». BMC Bioinformatics. 19 (1): 202. Дои:10.1186 / s12859-018-2187-1. ЧВК 5984344. PMID 29855387.
- ^ Консорциум GTEx; Лаборатория, Центр анализа данных и координации (Ldacc) - рабочая группа по анализу; Группы статистических методов - аналитическая рабочая группа; Расширение групп GTEx (eGTEx); Общий фонд NIH; NIH / NCI; NIH / NHGRI; NIH / NIMH; NIH / NIDA; Исходный сайт коллекции биологических образцов - NDRI; Исходный сайт коллекции биологических образцов - RPCI; Основной ресурс биопрепаратов - VARI; Репозиторий Brain Bank - Банк Brain Endowment Университета Майами; Leidos Biomedical - Управление проектами; Исследование ELSI; Интеграция и визуализация данных браузера генома - EBI; Интеграция и визуализация данных браузера генома - Институт геномики Ucsc, Калифорнийский университет в Санта-Круз; Ведущие аналитики; Лаборатория, Центр анализа данных и координации (Ldacc):.; Управление программами NIH; Сбор биопрепаратов; Патология; рабочая группа по рукописи eQTL; Battle, A .; Brown, C.D .; Engelhardt, B.E .; Монтгомери, С. Б. (12 октября 2017 г.). «Генетические эффекты на экспрессию генов в тканях человека» (PDF). Природа. 550 (7675): 204–213. Bibcode:2017Натура.550..204А. Дои:10.1038 / природа24277. ЧВК 5776756. PMID 29022597.
внешняя ссылка
- Функциональная геномика: введение в ресурсы EBI на Train OnLine
- Программа Европейского научного фонда по границам функциональной геномики
- MUGEN NoE - Интегрированная функциональная геномика в моделях мышей-мутантов
- Понимание природы: функциональная геномика
- Биоинформатика и функциональная геномика - Сайт-компаньон для Биоинформатика и функциональная геномика, 2-е изд.
- КОДИРОВАТЬ
- 4-я конференция Европейского научного фонда по функциональной геномике и болезням
| Геномика | |
|---|---|
| Биоинформатика | |
| Структурная биология | |
| Инструменты исследования | |
| Организации |
|
| |